
1. �͂��߂�
�@�זE�ɂ́A������̃T�C�N���t�C����(CyP);�זE���^��CyPA�Ɩ������^��CyPB ���������A�݂��̃A�~�m�_�z��ɂ͗ގ���������B����in vitro�̍y�f�����́A�|�� �y�v�`�h�̃v�������̃A�~�h�����̃V�X����g�����X�w�ِ̈����̊���(PPIase�� ��)�������B���グ��CyP�́A���ׂĂ̐����זE�ɑ�ʂɑ��݂���`�����ł���A ���������̈ێ��A�ۑS�ɕs���ȋ@�\���ʂ����ƍl������ɌW��炸�A���̐��� �w�I�Ȗ������S�͖������_�������Ă��Ȃ��Bin vitro�Ŏ��R�Ɍ��ɖ߂邢������ �`�������ACyPA�̕t���ɂ�胊�t�H�[���f�B���N�����i����邱�Ƃ���t�H�[�� �f�B���O�Ɋ֗^���邱�Ƃ���������O���[�v������B������CyP���������y��ۂ� �~���^���g��������ɐ��炷�鎖���ACyP���t�H�[���f�B���O�Ɋ֗^���Ȃ����� �������BCyP��in vitro�̓����ƌq����������́A�V���E�W���E�o�G�̎��זE�ŁA �T�C�N���t�B�����̃z�����O�ł���j�iA�̃~���^���g.�̎�́A���h�v�V�����\�� ����R1���A���ו�̕\�ʂɉ^�ꂸ�A���E�̂ɒ~�ς���B����R1�́A�z�[���f�B ���O�͊������Ă͂��邪�A�{������ׂ����ʂɉ^�ꂸ�A�j�iA�͗A���o�H�Ɋ֗^ ���A�t�H�[���f�B���O�ɂ͊֗^���Ȃ����Ƃ����ꂽ[1]�BLuban��́ACyPA���q �g�Ɖu�s�S�E�C���X(HIV-1)��Gag�O��`�����ɒ��ڌ������邪�ACyPB��HIV-2 ��simian immunodeficiency virus(SIV)��Gag�O��`�����Ɍ������邱�Ƃ���� [2-4]�B�X�ɁAHIV-1��p55gag�̃J�v�V�h�`�����̃v���������b�`�ȗ̈悪CyPA�Ƃ̌������x�z���邱�Ƃ������������B
�@�����זE�ɋ��ʂɑ��݂��A�������ێ��ɂ���ʂŕs���̐������ۂɂ����Ė{ ���I�ȓ��������̂Ƃ��ăZ���T�C�N���̐Î~(Go)�ɕۂߒ��ƑϔM��������� �ߒ�������B�M�҂́A�זE���^CyPA�͑O�҂̉ߒ��ɁA�������^CyPB�͌�҂� �ߒ��Ɋ֗^����̂ł͂Ȃ����Ƃ̗\���̂��ƌ�����i�߂Ă���B
�@�זE���A�ʏ�̏�Ԃ��炸�ꂽ���ɂ����ꂽ���A(�Ⴆ�A�זE���̑a������ �ς��A���x���㏸����)�^���p�N���̃��[�v�ɂ����Ă��̗��[�͌Œ肳��Ă��� ���c�\�����Ƃ�Ƃ����CyP�̃��K���h�������ʂɔF������ĕt���A���̍\�� ���ێ������B���̏ɖ߂��������K���h�������B���̂悤�ȋ@�\��CyP���� ���Ƃ���Ƃ���܂�CyP�Ɍ����������K���h������������Ȃ����Ƃ��������B
�@���łɕ����咰�ۂ̍זE���^�T�C�N���t�B����A(e-CyPA)�ƃg���y�v�`�h (suc-Ala-Pro-Ala-pNA)�̕����̂�1.8��\�̌����\��[5]�ƍy��ۂ̍זE���^�T �C�N���t�B����A(y-cyPA)��1.9A�����\���ɂ��Ă܂Ƃ߁AcyP�̃y�v�`�h�����T �C�g�ɕt���@�\�ɂ��ċc�_����B
2. �����ƕ��@
�@�咰�ۂ�e-CyPA�ƃg���y�v�`�h(Suc-Ala-Pro-Ala-pNA)�̕����̂̌����́A�g���y �v�`�h�����^�m�[���ɗn�����AO.5mM�`�����A6mM�g���y�v�`�h�A10%���^�m�[ ���A30%�O�a�������܂ރg���X���_�ɏՉt(PH8.O)��10��l�̃h���b�v��46%�O�a �������܂ރg���X���_�ɏՉt�ɑ���20����hanging drop���C�g�U�@�ɂ�蕽�t �ɂ��ē����B�팋���@�ōX�ɑ傫�������������B�g���y�v�`�h�̃V�X�̃v�������� ����͓��ɁA���^�m�[���Ɉ͂܂�Đ����͂���₷���A�܂��A�`�����̑a�����̗� ������^�m�[�����z�ʂ���̂Ő����͂���₷���Ȃ�A�g���y�v�`�h���`�����ɕt �����Ǝv����B
�@�����́A�Ε����n�ɑ����A��ԌQP212121,a=66.33��,b=68.23��,c=40.03��, Z=1,Dc=1.203g/cm3,Vm=2.43A3/Da,Vso1:49.4%�ł���B
�@���ꂼ��1mM HgCl2(18����)�AO.5mMK2PtCl4(3����)�ASmCl3�A�̖O�a�n�t (3����)���܂�55%�̗����n�t�ɕꌋ����Z������3��̏d���q�U���̂��B�� �����Əd���q�U���̂̔��ˋ��x�́A���G�l���M�[�����w�������̕��ˌ������(�g ����1.00��)�Ƃ��A�r�[�����C��BL6A�ɔ�����ꂽ�����q�p���C�Z���x���O�J�� ����p���Ď��W�����B�ʑ��͑��d���^�u���@�ɂ��PROTEIN�ʼn�͂��A2.5�� �\��Figure of Merit�́AO.73�ƂȂ����BIRIS���[�N�X�e�W������CHAIN��O�v���O �����ŕ��q�\�z���s�Ȃ��AXPLOR�v���O�����ɂ�萸�������ŏI�I�ɁA5.0����1.8 ����12783�̓Ɨ��Ȕ���(����\�Ȕ��ː���79%)�ɑ�R=17.6%�Ɏ��������B �y��ۂ�y-CyPA�̌����́A�|���G�`�����O���R�[���𒾓a�܂Ƃ����g���X���_�� �Չt(PH8.O)��p���āA20����hanging drop���C�g�U�@�ɂ�蓾�A�X�Ɏ팋���@�� ��茋����傫�������������B
�@�����́A�O�Ώ��n�ɑ����A��ԌQP1, a=44.45(2)��, b=53.11(2)��, c=32.018(6)��, ��=84.91(2)߁A��=95.22(4)�,��=108.56(4)�,Z=2,Dc=1.21g/cm3,Vm=2.07A3/Da,Vsol=40.6%�ł���B
�@�ꌋ���̔��ˋ��x�̎��W�́A���ˌ�������Ƃ�65303�̊ϑ��f�[�^�ɑ� Rmerge=4.7%�ƂȂ����B�q�g�זE���^�T�C�N���t�C����(h-CyPA)161���T�[�`���f��. �Ƃ��ĕ��q�u���@�ɂ��XPLOR�v���O�����ʼn�͂������ʁA�œK��2���q�� rigid-body�̐�������R���q=34%��^�����BO�ŕ��q�\�z���s�Ȃ��AXPLOR�ɂ�� �������́A�ŏI�I�ɁA5����1.9����17931��(����\�Ȕ��ː���85.8%)�̓Ɨ��� ���˂ɑ�R=17.6%�Ɛ������B
3. �זE���^�T�C�N���t�C����A(e-CyPA,y-CyPA)�̑S�̍\��
�@3�����\������ɕ��ׂ��咰�ہA�y��ہA�q�g�R���זE��CyPA(e-CyPA, y-CyPA,h-CyPA)�Ɩ������^CyPB(e-CyPB,y-CyPB,h-CyPB)�̃A�~�m�_�z���} 1�Ɏ����B�q�gh-CyPA�́A5�̃G�L�\����4�̃C���g�������琬��Bh-CyPA �Ƃ̑������́A�M���ނł�95%�ȏ�ƍ����Ay-CyPB�Ƃ�60%�ŁAe-CyPA�ł�27% �ɕۂ���Ă���B�זE��CyPA�Ɩ������^CyPB�̑������́A�咰�ۂ�54%,�y��� 56%,�q�g65%�ł���B
�}1.�q�g�A�y��ہA�咰�ۂ̍זE���^CyPA�Ɩ������^CyPB�̃A�~�m�_�z��B �n�C�t���͌����A�e�����̓��K���h�����T�C�g�A�������͓����A�~�m�_�ł��邱 �Ƃ������B����h-CyPA�̃G�L�\���̋��E�ł���B
e-CyPA��y-CyPA�̑S�̂̍\����(�}2)�A1���̃A�~�m�_�z��\�z������ ��A�\���̍��i�͓����ł���B���҂Ƃ�8�{�̋t���s�̃��X�g�����h�����o�[���� �\�����\�����A���o�[�����̊W������悤�ɏ㉺�����w���N�X����߂�B���o�[�� ���́A4�{�̋t���s�̃��X�g�����h���琬��2���̃��V�[�g���������ďd�Ȃ�B�� �E���^h-CyPA161�Ɩ������^h-CyPB171�̑S�̂̍\���Ƃ������ł���B
�@�����w�I�ɓƗ���2�̕��qy-CyPA�Ԃ̃A�~�m�_�c��C�����d�˂�����C�������q�Ԃ̂���̕���r.m.s.�́A0.25���ł���Ay-cyPA��h-cyPA�̑Ή�����A�~�m�_�c��Ƃ̂���́A0.38A�ł���3�����\�������S�ɕۑ�����Ă���B
�@e-CyPA�́Ay-CyPA�̍��i���Ȃ�8�{�̃��X�g�����h��2�{�̃��w���b�N�X���\�� ����75�̃A�~�m�_�c���C���̌��q�Ԃ̂���̕��ς́A0.67���ŗǂ��d�Ȃ��Ă� ��A������{�I�\��������(�}3)�B
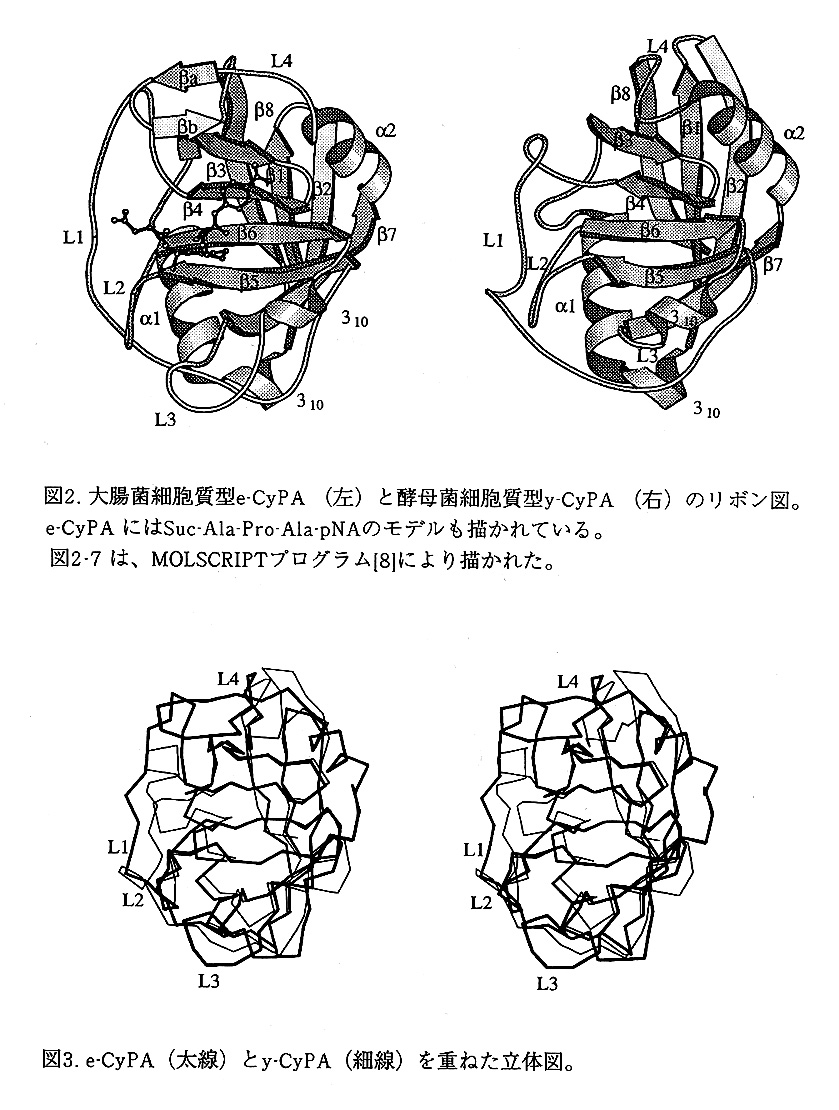
�@e-CyPA,y-CyPA,h-CyPA�́A���ׂď㑤�̃��V�[�g(��3,��5,��6,��5�X�g�����h) �̕\�ʂ́A�y�v�`�h�̌����T�C�g�����B����A�����̃��V�[�g(��8,��1,��2,��7 �X�g�����h)�́A�㑤�̃��V�[�g�Ƃ̑a�������ݍ�p�ɂ����o�[�����\�������艻 ������B�X�ɁA2�{�̃��w���N�X���S�̂̍\���̈��艻��⋭����B
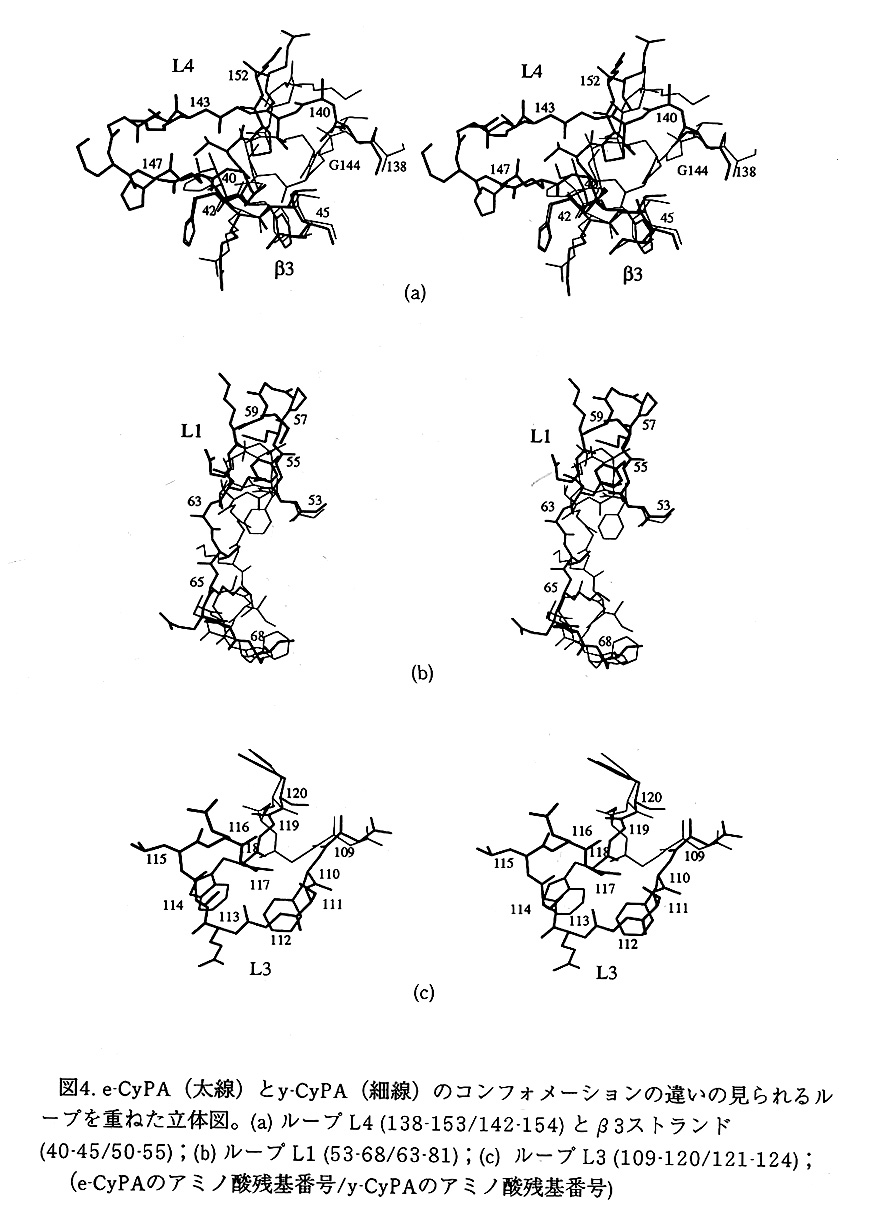
�@e-CyPA�́A�㑤�̃��V�[�g�̎��ӂɂ���L1,L3,L4�̃��[�v�ƃ�1�ƃ�3�����ԃ� �[�v�̃R���t�H���[�V�����ɂ�����y-CyPA,h-CyPA�ɑ��Ⴂ��������(�}4)�B e-CyPA��L3��L4���[�v�ɂ�����A�~�m�_�z��̑}����L1���[�v�ɂ����錇�����R ���t�H���[�V�����ω��������炷�B�܂��Ae-CyPA�ɂ����āA��1�ƃ�2�̃X�g�����h ���Z���̂͌����ɂ�萶����Be-CyPA�̃�2�ɑ���L4���[�v�́A��3�X�g�����h���� �����������R�[�X��ʂ��ă^�[���������{�������B���̃R���t�H���[�V�����́A �q�g�������^h-CyPB�ɂ�������Be-CyPA��Ala140�ɑΉ�����ʒu��y-CyPA, h-CyPA�ł́A�����̂Ȃ��O���V��������(���ꂼ��Gly144,Gly146)�A��3�Ɨ��̏�Q ���Ȃ��Bh-CyPB�ł͑����̂��郊�W������߁AL4���[�v�́Ae-CyPA�Ɏ����R ���t�H���[�V���������Be-CyPA��L1���[�v�́AL4���[�v�̃����{����His147�A ��3��Ile40, His42��Phe55, Pro57, Met59�Ƃ̑a�����̑��ݍ�p��ʂ��Ė��Ƀp�b �L���O����悤�ȃR���t�H���[�V�������Ƃ�BSer111����Trp118��8�̃A�~�m �_�c��̑}��������e-CyPA��L3���[�v�́A���̕����݂̂q�̕\�ʂɓ˂��o���� ����B
4.�y�v�`�h�����T�C�g
�@e-CyPA,y-CyPA,h-CyPA�́A���ׂď㑤�̃��V�[�g�̕\�ʂɃ��X�g�����h�ɕ��s
�ɑ���a������B���̍a�ɂ́A�a�����̃A�~�m�_�c��ň͂܂ꂽ�|�P�b�g������B
y-CyPA�ɂ����ẮA5�̃A�~�m�_�c��̑a�����̑���(Phe58,
Met59, Trp119, Leu120, His124)���|�P�b�g�̕ǂ��`�����A���Phe111����߂�(�}5)�BPhe58,
Met59�́A��3�ƃ�4�̃^�C�vII�̃��^�[���Ɉʒu���A������Phe58,
Met59�̓��o�� �W�l�\���������A���V�[�g�̓������������B�܂��A������Trp119,
Leu120 ��1�^ �[��310�w���b�N�X��C�����߂�BLeu120�̎卽�̃J���{�j����́A��7�X�g
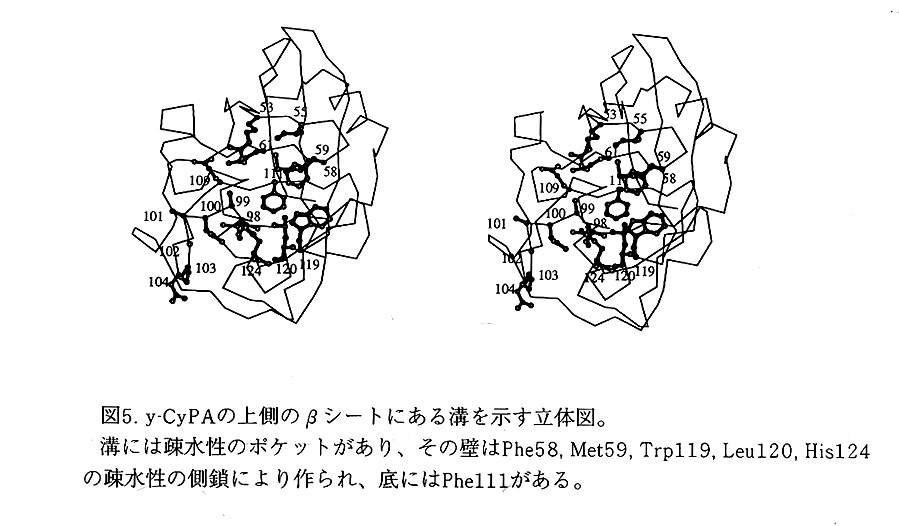
�@�����h��1�O�ɂ���His124�̎卽�̃A�~�h��NH�Ɛ��f���������BHis124�� ������NH�́AMet98�̎卽�̃J���{�j����Ɛ��f���������A�����̖F�����\�� �ɑ������ɌŒ肳���BPhe58, Trp119, Phe111�̑����̖F���́A�`�����̕\�� �ɕ��s�Ȕz�������Bh-CyPA�ɂ��������̑a�����̃|�P�b�g�́A�����A�~�m�_�c�� (Phe60, Met61, Trp121, Leu122, His126, Phe113)�������ʒu���߁A�����z�� �����B
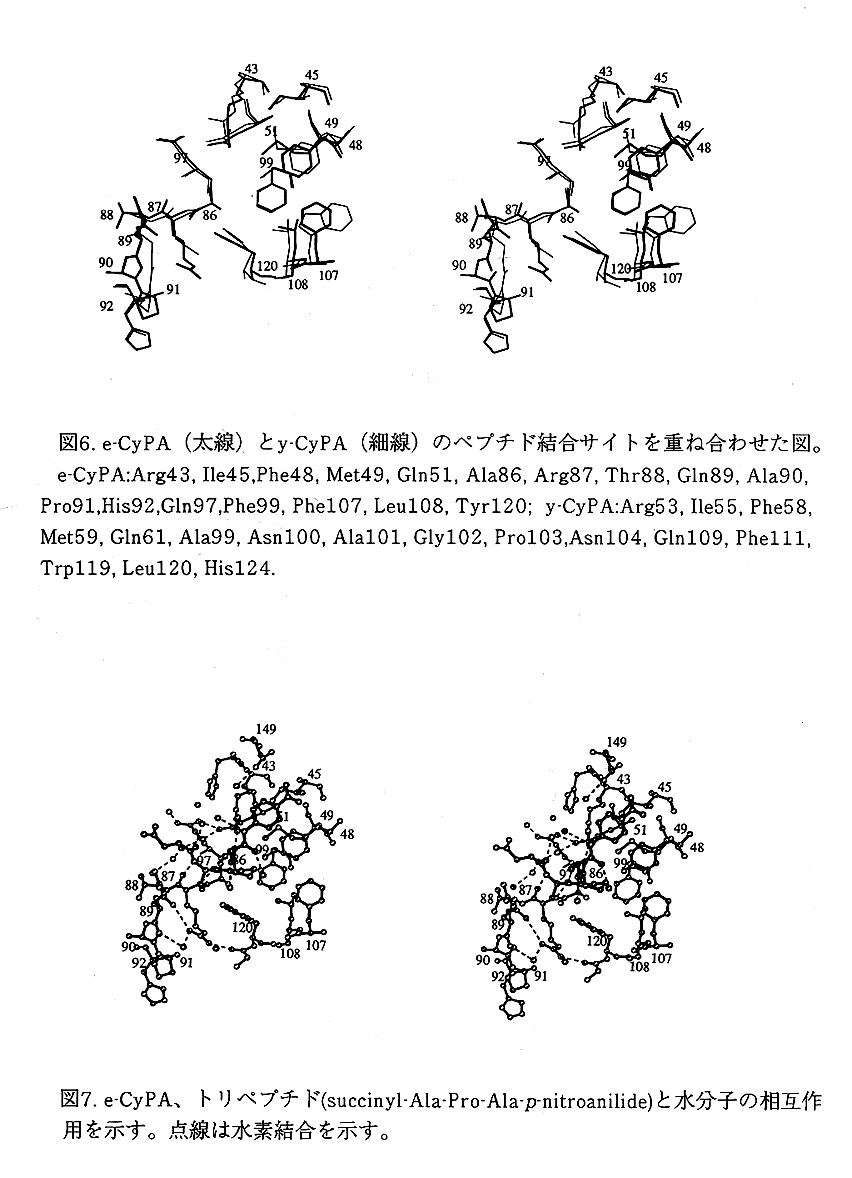
�@e-CyPA�ł��A�㑤�̃��V�[�g�̕\�ʂ̍a�ɑa�����̃|�P�b�g������B���l��5�� �̃A�~�m�_�c��̑a�����̑���(Phe48, Met49, Phe107, Leul08, Tyr120)���ǂ��� ��APhe99����ɂ���(�}6)�BPhe48��Met49�́A���o���W�l�\���������A���V�[ �g�̓������������APhe107��Leu108��1�^�[��310�w���b�N�X��C�����߂�B Tyr120�̎卽�̃A�~�h��NH�́ALeu108�̎卽�̃J���{�j����Ɛ��f���������� �艻����Ă���BTyr120�̑����̖F���́A�t�F�j����̐��_���n�}�ɎN���悤 �ɕ��q�ʂɑ������ɗ����オ��B����APhe48, Phe107, Phe99 �̑����̖F���� ���q�ʂɕ��s�ɕ��ԁB
�@�^�j�����ŕۑ�����Ă���g���v�g�t�@���ƃq�X�`�W�����咰�ۂł̓t�F�j���A ���j���ƃ`���V���ɕψق��Ă��邯��ǂ��A`�a�����|�P�b�g���`������A�~�m�_�c ��̑����́A���ׂē����z�������B�����̃A�~�m�_�c��͍y��ۂ���q�g�Ɏ� �邷�ׂĂ̐^�j������CyPA�ŕۑ�����Ă��邱�Ƃ��A�����̔z���́A���j���� �Ɛ^�j�����̂��ׂĂ�CyPA�ŕۑ�����Ă���Ǝv����B
5.�g���y�v�`�h�Ƒa�����̃|�P�b�g�̑��ݍ�p
�@e-CyPA�ƃg���y�v�`�h(Suc-Ala-Pro-Ala-pNA)�̕����̂̍\���ɂ����āA�g���y�v �`�h�̓V�X�ِ��̂̌`�Ńv��������5����Tyr120�̑����ɕ��s�ɂ��đa�����̃| �P�b�g�ɑ}�����ĕt���Ă���(�}7)�BArg43��N��2�́A�g���y�v�`�h��Pro2�̎� ���̃J���{�j����Ɛ��f���������BArg43��N��1�́APro2�̎卽��C=O���܂��� ���L����O����3.21���Ɉʒu���AAsp149�̑����ɐ��f�������鐅���q����3.31�� ����Ă���BArg43��N�ẤAGln51��O��1����3.09���ɂ���BArg43�̑����̃O �A�j�f�B���́AIle45��Phe48�̑a�����̑����ƃg���y�v�`�h�̃A�j�����Ɉ͂� ��Ă���A�C�I��������Ă��炸�A�d�דI�ɒ����ł���BArg43,Gln51,Gln97�� �����́A���f�����̃l�b�g���[�N������B���̃l�b�g���[�N�́A86����88�̎c ��̎卽�ƕ��s�ɑ���A�g���y�v�`�h�̃v��������N�đ�����߂�a�����B���� Arg87�̎卽���g���y�v�`�h��Ala1�̎卽�Ƌt���s���V�[�g�̂悤�ɐ��f�������� ��B�c��̃g���y�v�`�h�̎卽�́A4�̐����q�Ɛ��f�������`������BPro2�̃A �~�h�̒��f���q�݂̂��`�����Ɩ{�f��������炸�ATyr120�̖F����Gln51�̑��� �̒��Ԃɂ���B
�@y-CyPA�́A���݂̂Ƃ���y�v�`�h�Ƃ̕����̂̌����͓����Ȃ��B�T�C�N���X�| ����A(CsA)�̕����̂̌����́A5���̕���\�œ�����͂����B������5�ʑ̂� �`������By-CyPA�P�Ƃ̌����ɂ�2���q���݂��AArg53�̃O�A�j�f�B����̌����� �قȂ�Be-CyPA��Gln51��Gln97�ɑΉ�����y-CyPA��Gln61��Gln109�́A�����z �������Đ��f�������A99����101�̎c��̎卽�����s�ɑ���A�y�v�`�h�̃v������ ��N�đ�����߂�a�����B
�@h-CyPA�ƃe�g���y�v�`�h(acetyl-Ala-Ala-cis-Pro-Ala-amidomethylcoumarin)�̕� ���̂̍\����͂̌���1g1�́Ae-cyPA�Ɠ��l�AArg55, G1n63, Gln111�̑����́A���f �����̃l�b�g���[�N������A101����103�̎c��̎卽�����s�ɑ���A�e�g���g ���y�v�`�h�̃v��������N�đ������̍a���߂�BAsn102�̎卽���e�g���y�v�` �h��Ala2�̎卽�Ƌt���s���V�[�g�̂悤�ɐ��f���������B�����̃A�~�m�_�c�� �́A���ׂčy��ۂɂ����Ă��ۑ�����Ă���B
�@e-CyPA�ƌ�������g���y�v�`�h�̃�/�Ղ�2�ʑ̊p�́AAla1,Pro2,Ala3�ɑ� �Ă��ꂼ��-119/146�,-69/143�,-67/146�,�ł���B�g���y�v�`�h��C���� p-nitoani1ide�́APhe48�Ɨאڂ��镪�q��Pro67�Ƒ��ݍ�p����Bh-CyPA�ł́A�e�g ���y�v�`�h��coumarin���APhe58�Ɨאڂ��镪�q��Trp121�Ƒ��ݍ�p����B���̕� �q�Ԃ̑��ݍ�p���Ah-CyPA�ƌ�������e�g���y�v�`�h�Ƃ́A�v��������C�đ���2 �ʑ̊p�̈Ⴂ�݁APro3��A1a4�̃�/�Ղ�2�ʑ̊p�́A���ꂼ�� -72/-179�,-123/71߂ł���B
�@e-CyPA�ƌ�������g���y�v�`�h��Ala-Pro�A�~�h�����̃֊p�x�́A-1.2߂ŁA�c�C�X �g�t�H�[����90߂���傫������Ă���B
6. �l�@
6.1 �g�����X�ِ��̂�CyPA�̃g���y�v�`�h�����T�C�g�ɂ���ĔF������Ȃ�
�@�����{�k�N���A�[�[A�A�Y�_�E���f�y�fII�A�R���[�Q���^�C�vIII�A���{�k�N���A�[ �[T1���ϐ��܂Ńt�H�[���g�������ꂽ��A�����̕ϐ��܂𔖂߂�Ƃ����̒`�� ���͎��R�Ɍ��ɖ߂邪�A���t�H�[���h�̉ߒ���CyP��������Ƃ������i�ޒi�K�� ��������邱�Ƃ����ꂽ�B�����̒`�����̃V�X�[�v�������̃Ղ�2�ʑ̊p�́A e-CyPA�ƌ�������g���y�v�`�h�̂����=143߂Ƃقړ����p�x�����B�����A���� �̃V�X�[�v�����������`����(�T�[�����C�V���A�T�u�`���V���A�G���X���L���� �����A�u�h�E���ۂ̃k�N���A�[�[��)�ł́A�V�X�[�v�������̃J���{�j����CO���A �t����������CyP�ɂ���ĉ�������Ȃ��B���̂悤�ȋt�̔z���́ACyP�߂Â��� (�v��������)�ɐ�������ɕt����CyP�����T�C�g�ɋߕt���Ȃ�����ł���B �V�X�[�g�����X�ِ��������̃A�b�Z�C�ŁALiCl���܂ރg���v���I���G�^�m�[���n �t�ɗn������Suc-Ala-Ala-(cis-Pro)-Phe-PNA�̃~�J�G���萔Km�́Ah-CyPA�ɑ��� O.87mM�ł���B�����A�b�Z�C�ŁADMSO�ɗn��������ł���v���������܂ރy�v �`�h��Suc-Ala-Ala-(cis-Pro)-Phe-pNA�̑���ɑ��āA�����T�C�g�ɋ������ē����� ��Kd�́A20mM�ȏ�ƌ��ς�ꂽ�B�����̃y�v�`�h�́A�܂���ł��肻��Km�� Kd�Ɠ����ł���B�V�X�ƃg�����X�ِ̈��̂̍������̔䂪O.1�ł���̂ŁA�V�X�[�v �������̃y�v�`�h��Km�́A2mM�ȉ��ł���A����g�����X�ِ��̂�Km�́A20mM �ȉ��łȂ����Ƃ������B�X�ɁASuc-Ala-Leu-(cis-Pro)-Phe-pNA�̃V�X�[�g�����X�ِ� �������A�b�Z�C�ɂ����ē�����PPIase����������FKBP�ɑ���~�J�G���萔 Km=0.5mM�ɑ��ASuc-Ala-Leu-Pro-phe(cis/trans=1/9)��������������Kd�́A 3mM�ł���B���̃y�v�`�h��Km�́AKd�Ɠ�������B�]���āA�g�����X�[�ِ��̂� Km�́A�ق�30mM�ł��邱�Ƃ������B�����̂��Ƃ́ACyPA��FKBP���g�����X�� ��V�X�w�ِ̈��������ɂ͓����Ȃ����Ƃ��Î�����B���Ȃ킿�V�X����͂��ɘc�� ���t�H�[�����g�����X����傫�����ꂽ�t�H�[�������̑a�����̃|�P�b�g���F���� �邱�Ƃ������B�X�Ɍ����ACyPA�̑a�����̃|�P�b�g���`������5�̃A�~�m�_�c ��́A�c�V�X�ƃc�C�X�g�t�H�[���������̃|�P�b�g�ɗU�����Ȃ��B
6.2 ���K���h�̌����@�\
�@�g���y�v�`�h(Ala-Pro-Ala)�́A�v���������V�X�^��������ꍇ�A���Ƃ̑��ݍ�p ���l������Ɨ�����Ala�݂̂ŒZ���Ƃ���炪�c�R���t�H���[�V����������Ă� �G�l���M�[�I�ɑ傫�ȑ��ɂȂ炸�A����V���^�̓��ʂȃR���t�H���[�V������ CyPA�̃y�v�`�h�����T�C�g�Ƀt�B�b�g����BSuc-Ala-Leu-cls-Pro-Phe-pNA���邢�� �����蒷���y�v�`�h���ł́A�S�̂ŏ����c��Ŋɘa���A�v�������̃V�X�^�� �c�C�X�g�����ɂ���B�]���āA������c�C�X�g�^�̃V�X����g�����X�w�ِ������� ���ACyP�ɂ���Ă��̎��͂̐��������������ȃg�����X�w��������A���̏�Q �ŗ���Ă����BCyP�Ɍ�������V�X�^���Ƃ�ƃv�������̐�̃A�~�m�_��CyP�Ƃ� ���藣���BCsA�ɂ����Ă͊���邱�Ƃɂ��V�X�^���n�t�ő��݂ł��邪�A CyP�ɔF������đa�����̊��ɗ���Ǝ��͂̐��������������ȃg�����X�w�� �������傫�ȃR���t�H���[�V�����ω����B�y�v�`�h�̃v���������Ae-CyPA�ƌ� ������g���y�v�`�h�̃V�X�[�v��������2�ʑ̊p��=143߂���������A���M�j���� ����(Arg43(e-CyPA);Arg53(y-CyPA))�Ɛ��f���������y�v�`�h�����T�C�g�ɕt���� �Ƃ��o����B�������A��=143߂̔z���́A�����y�v�`�h���邢�͒`�����̃��[�v�� �͕s����łقƂ�ǎ��Ȃ��B���̔z���������������łƂ��g���y�v�`�h�́A�� ��C=O��Arg43�̑����Ɛ��f������������艻���Ă��������Ǝv����B
�@HIV-1��Gag�̃J�v�V�h�`�����́ACyPA��Pro222�ŕt�����ACyPB�ɂ͕t���Ȃ��B �����AHIV-2�̂����CyPA�ɕt����CyPB�ɕt���B�X�ɁACyPB�́Asimian immunodeficiency virus(SIV)�ł���SIVMAC239'SIVAGMSab384�ɕt���BCyPA��CyPB�̃y�v�`�h�����T�C�g�͂قڕۑ�����Ă��邪�A�y�v�`�h�����T�C�g�̋ߖT�ōזE���^CyPA�َ̈�ԂƖ������^CyPB�َ̈�Ԃł́A���ꂼ��ɋ��ʂł��邪CyPA��CyPB�Ԃł͈قȂ�̈悪L2�̃��[�v�̃^�[���Ɍ�����B���̃��[�v��N�đ��́A �y�v�`�h��N�đ��̎卽�Ɛ��f�������B�זE���^y-CyPA��h-CyPA�ł� NAGPNTNGS�ł���A�������^��y-CyPB��h-CyPB�ł�NA/RGKDTNGS�ŁACyPA�� �Œ�^�v�������[�A�X�p���M���ɑ�CyPB�̓C�I�����c��̃��W���[�A�X�p���M�� �_�ł���B�咰�ۂł�1�c��̑}�������邪�Ae-CyPA�ł́ARTQAPHSATA�ł��� ����AeCyPB�ł�RTADKDSATS�őO�҂͌Œ�^�v�������[�q�X�`�W���A��҂́A�C �I���^���W���[�A�X�p���M���_�ł���B���̋͂��ȈႢ���AGag�̃J�v�V�h�`���� �̋͂��ȈႢ����ʂ��A�X�ɂ́A�זE���^CyPA�Ɩ������^CyPB�ɕt�����K���h�� ��ʂ���Ǝv����B����A�������^e-CyPB,y-CyPB�ɂ��ă��K���h�������ʂ� �ڍׂȍ\���ׁACyPA��CyPB�̃y�v�`�h�����@�\�ɂ����͂��ȍ���������� ���Ƃɂ��A�����w�I�@�\��̈Ⴂ���𖾂��邱�Ƃ��\�ƍl����B
�Q�l����
[1] N. .J. Colley, E: K. Baker, M. A. Stamnes, C. S. Zuker, Cell, 67, 255-263 (1991).
[2] J. Luban, K. L .Bossolt, E. K. Franke, G.V. Kalpana, S.P.Goff, Cell, 73,
1067-1078 (1993).
[3] E. K. Franke, H. E. H. Yuan, J. Luban, Nature, 372, 359-362 (1994).
[4] M.Thali, et al., Nature, 3 72, 363-365 (1994).
[5] M. Konno, M. Ito, T. Hayano, N. Takahashi, J. Mol. Biol. 256, 897-908 (1996).
[6] H. Ke, J. Mol. Biol. 2 28, 539-550 (1992).
[7] V. Mikol, J. Kallen, G. Pflugl, M. D. Walkinshaw, J. Mol. Biol. 234, 1119-1130, (1993).
[8] P. J. Kraulis, J. Appl. Crysttalog., 24, 946-950 (1991).
[9] J. Kallen, M. D. Walkinshaw, FEBS Letters, 300, 286-290 (1992).