
偼偠傔偵
兛亅傾儈儔乕僛偼丄揵暡傗條乆側僆儕僑摐偺兛-1丆4僌儖僐僔僪寢崌傪壛悈暘夝偡傞峺慺偱偁傞丅偙偺峺慺偼丄岺嬈梡峺慺偲偟偰尰嵼嵟傕戝検偵巊傢傟偰偄傞峺慺偱偁傝丄庡偵丄揵暡偐傜偺摐惢憿偵偍偄偰揵暡傪嵟弶偵塼壔偡傞岺掱傗丄晍傪怐傞帪偵巺偺妸傝傗嫮搙傪崅傔傞栚揑偱壛偊傜傟傞揵暡屝傪彍嫀偡傞岺掱偱戝検偵巊傢傟偰偄傞丅偙偆偟偨岺掱偱梡偄傜傟傞峺慺偼丄偦偺傎偲傫偳偑 Bacillus 懏桼棃偺傕偺偱偁傞丅偙傟偼丄旝惗暔桼棃偱偁傞偨傔戝検惗嶻偵岦偄偰偄傞偙偲偵壛偊偰丄 Bacillus 懏桼棃偺峺慺偑桪傟偨懴擬惈傪帵偡偙偲偵傛傞丅
丂変乆偼丄 Bacillus 懏桼棃偺兛亅傾儈儔乕僛偺懴擬惈偲暘巕峔憿偲偺娭楢惈偵嫽枴傪帩偪丄拞掱搙偺懴擬惈傪帵偡B.amyloliquefaciens 桼棃偺兛亅傾儈儔乕僛(帄揔壏搙55-60亷丄埲壓BAA偲棯偡)偲崅偄懴擬惈傪帵偡B.lichenifoformis 桼棃偺兛亅傾儈儔乕僛(帄揔壏搙80-85亷丄埲壓BLA偲棯偡)偺2偮偺峺慺偺X慄寢徎夝愅傪峴偄丄椉幰偺棫懱峔憿偺堘偄偐傜峺慺偺懴擬惈偺儊僇僯僘儉傪柧傜偐偵偟傛偆偲峫偊偨丅BAA偲BLA偼嫟偵丄傾儈僲巁巆婎悢483丄暘巕検5,5000偱偁傝丄傾儈僲巁攝楍偺摨堦惈傕80%偲崅偄丅傑偨丄椉峺慺偲傕妶惈偺敪尰偵偼僇儖僔僂儉僀僆儞偑昁梫偱偁傞丅偙偙偱偼丄BAA偲BLA偺2偮偺峺慺偺寢徎夝愅偺夁掱偲丄椉幰偺峔憿偺斾妑偺寢壥柧傜偐偵側偭偨BLA偺懴擬壔偺婡峔偵偮偄偰弎傋傞丅
幚尡
BAA偲BLA偺寢徎夝愅
丂椉峺慺偺寢徎夝愅偼1989擭偵摨帪偵巒傔傜傟偨偑丄屻弎偡傞傛偆偵側偐側偐椙偄寢徎偑摼傜傟偢丄夝愅偑愭偵恑傫偩BLA偵偮偄偰暘巕儌僨儖偺慻傒棫偰偑壜擻側揹巕枾搙恾偑摼傜傟偨偺偼丄1995擭偱偁偭偨丅偟偐偟側偑傜丄偙偺擭偵Huber傜偺僌儖乕僾偵傛偭偰BLA偺寢徎夝愅1)偑丄変乆偺傕偺偲偼堎側傞徎宯偱峴傢傟偰偟傑偭偨丅偦偙偱丄偙偺擭埲崀偼丄BAA偺夝愅偑庡偲側偭偨丅
丂BAA偺夝愅偼丄BLA偺尨巕嵗昗偑擖庤偱偒側偐偭偨偨傔丄変乆偺僌儖乕僾偱MlR朄+density modification 偵傛傝摼傜傟偨BLA偺揹巕枾搙暘晍傪儌僨儖暘巕偲偟偰暘巕抲姺朄偵傛傝峴偭偨丅側偍丄BLA偺夝愅傕変乆偺徎宯偺傕偺偵偮偄偰惛枾壔傑偱峴偄丄BAA偲偺峔憿偺斾妑偵梡偄偨丅
寢徎壔偺挿偄摴偺傝
丂BAA偺嵟弶偺寢徎壔僗僋儕乕僯儞僌偱摼傜傟偨寢徎偼旕忢偵嵶偄恓忬徎(懢偝0.01mm埲壓)偱偙傟傪戝偒偔偡傞偙偲偑傑偢戝曄偩偭偨丅偟偐偟側偑傜丄抈敀幙擹搙傪嬼慠0.5%偐傜1%偵忋偘偨偙偲偵傛傝丄恓忬徎偼暯峴榋柺懱忬偺奜尒偺寢徎偵曄 傢傝丄僒僀僘偺栤戣偼夝寛偝傟偨2)丅偟偐偟側偑傜丄摨偠傛偆側暯峴榋柺懱忬偺寢徎偱傕丄扨幬徎偲幬曽徎偺2偮徎宯偑偁傞偙偲偑夝偭偨丅夝愅偵偼旕懳徧扨埵拞偵娷傑傟傞暘巕悢偺彫偝偄幬曽徎宯偺寢徎傪慖傫偩偑丄奜尒偐傜尒暘偗傞偙偲偺偱偒側偄懠曽偺寢徎偺懚嵼偵偢偭偲擸傑偝傟偨丅傑偨丄偙偺寢徎偺惉挿丒曐懚偺偨傔偵偼儕儞巁娚徴塼偑昁恵偱丄偙傟偑BAA偱廳尨巕桿摫懱偑尒偮偐傜側偐偭偨庡場偲峫偊傜傟傞丅
丂BLA偺嵟弶偺寢徎壔僗僋儕乕僯儞僌偱偼丄敄偄斅忬徎偑岦偒傪曄偊側偑傜懡悢愊憌偟偨寢徎偑摼傜傟偨丅偙偺寢徎偼丄pH丄抈敀幙擹搙丄捑揳嵻擹搙丄寢徎壔壏搙摍偺挷愡偱偼扨寢徎偵側傜側偐偭偨丅偦偙偱丄寢徎曣塼偵庬乆偺揧壛暔幙傪彮検壛偊傞幚尡傪峴偭偨丅偦偺寢壥丄尒偮偐偭偨偺偑EDTA偱偁傞丅兛-傾儈儔乕僛偼丄峺慺妶惈傪曐偮偨傔偵僇儖僔僂儉傪昁梫偲偡傞偨傔丄僇儖僔僂儉傪峺慺偐傜扗偄庢傞EDTA偺揧壛偼丄暘巕偺棫懱峔憿傪夡偡婋尟惈偑偁傞丅偟偐偟側偑傜丄摉帪戝妛堾惗偩偭偨埳摗壝復巵偼丄偁偊偰EDTA傪揧壛偟丄偒傟偄側拰忬徎傪摼傞偙偲偵惉岟偟偨2)丅
丂BLA偱偼偦偺屻丄巊梡偱偒傞廳尨巕壔崌暔偺庬椶傪憹傗偡偨傔偵寢徎壔PH傪拞惈偵偡傞丄媦傃丄摨宆抲姺朄傪峴偆偺偵廫暘側惛搙偺夞愜僨乕僞傪廂廤偱偒傞寢徎僒僀僘傪妋曐偡傞偨傔丄偺寁2夞偵傢偨傝寢徎壔忦審偺曄峏偑峴傢傟偨丅偙傟傜偼丄寢徎偺栤戣偱廳尨巕桿摫懱偺扵嶕偑峴偒媗傑偭偨偙偲偵婥偯偔搙偵峴傢傟偨偺偱丄寢壥偲偟偰寢徎夝愅偑戝偒偔抶傟偰偟傑偭偨丅椉傾儈儔乕僛偵偍偄偰嵟廔揑偵寢徎夝愅偵梡偄偨寢徎偺寢徎壔忦審媦傃寢徎妛揑僨乕僞傪昞1偵帵偡丅側偍丄夝愅偺峴偊傞寢徎偑摼傜傟傞傑偱偵丄BAA偱偼2擭偺嵨寧偑丄偦偟偰BLA偱偼幚偵6擭偺嵨寧偑棳傟偨丅
曻幩岝X慄傪梡偄偨夞愜嫮搙僨乕僞偺廂廤
丂BAA丒BLA嫟丄X慄夞愜嫮搙僨乕僞偺廂廤偼丄慡偰崅岺尋曻幩岝巤愝偺BL-6A2價乕儉儔僀儞偵偍偄偰抈敀幙梡儚僀僙儞儀儖僌僇儊儔偲僀儊乕僕儞僌僾儗乕僩傪梡偄偰峴偭偨丅夞愜僀儊乕僕偺張棟偵偼丄僾儘僌儔儉WEIS傪梡偄偨丅BAA偺Native偺夞愜僨乕僞丄媦傃丄BLA偺Native偲3偮偺廳尨巕桿摫懱偐傜偺僨乕僞廂廤偺寢壥傪昞2偵傑偲傔傞丅
BLA偵偍偄偰丄Native偲奺桿摫懱娫偺夞愜嫮搙偺嵎偼10%慜屻偲彫偝偔丄僔儞僋儘僩儘儞曻幩岝傪巊偭偰應掕岆嵎傪彫偝偔梷偊傞偙偲偑昁恵偱偁偭偨丅
MIR朄偵傛傞BLA偺弶婜埵憡偺寁嶼
丂BLA偱偼丄Sm丄Pt丄Hg偺3偮偺廳尨巕桿摫懱偑摼傜傟丄MIR朄偵傛傝埵憡傪寁嶼偡傞偙偲偑偱偒偨丅廳尨巕僷儔儊乕僞乕偺惛枾壔偲埵憡偺寁嶼偵偼丄CCP4偺MLPAPE傪梡偄偨丅3偮偺廳尨巕桿摫懱偲傕堎忢暘嶶偺岠壥傕娷傔偰埵憡傪寁嶼偟偨偑丄10乣3侌暘夝擻偵偍偗傞暯嬒偺Figure of merit偼0.39偱偁偭偨丅摼傜傟偨揹巕枾搙恾偼丄5侌暘夝擻偱偼丄暘巕椞堟偑堦栚偱夝傞椙幙偺傕偺偱偁偭偨偑丄3侌暘夝擻偱偼丄僲僀僘偑懡偔丄1杮偺兛僿儕僢僋僗偑丄偐傠偆偠偰僩儗乕僗壜擻側掱搙偱偁偭偨丅偦偙偱丄CCP4偺DM傪梡偄偰揹巕枾搙恾偺夵慞傪峴偭偨丅
DM偵傛傞BLA偺揹巕枾搙恾偺夵慞
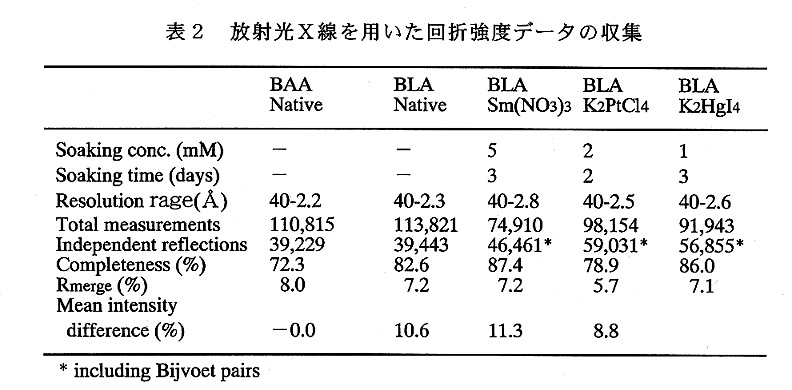
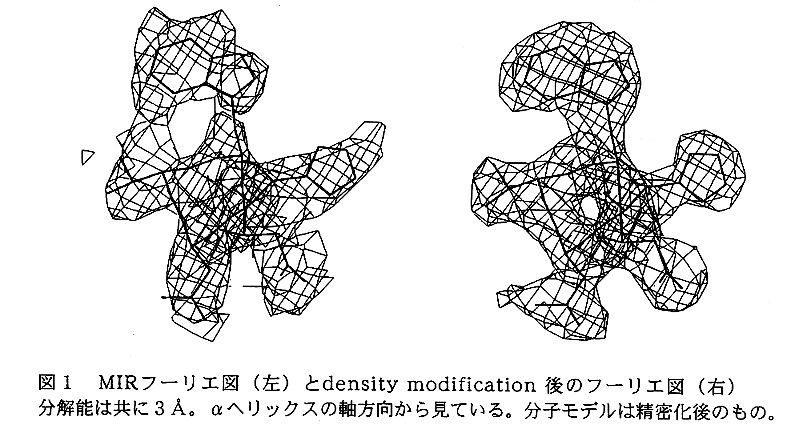
丂Non-Crystallographic Symmetry averaging(NCS averaging)傪岠壥揑偵峴偆偨傔偵偼丄1)暯嬒壔偑峴傢傟傞斖埻=暘巕椞堟傪惓妋偵寛掕偡傞偙偲丄2)旕寢徎妛揑懳徧(NCS)傪婰弎偡傞夞揮丒暲恑僷儔儊乕僞乕傪惓妋偵媮傔傞偙偲偑昁梫偱偁傞丅BLA偺夝愅偱偼丄1)傪峴偆偨傔偵暘巕儅僗僋嶌惉巟墖僾儘僌儔儉僷僢働乕僕MASKMAKE偑怷晹偵傛傝奐敪偝傟偨丅偙偺僾儘僌儔儉偼丄僔儕僐儞僌儔僼僀僢僋僗幮偺IRIS僔儕乕僘梡偵奐敪偝傟偨傕偺偱丄僨傿僗僾儗僀忋偵昞帵偝傟偨揹巕枾搙恾傪尒側偑傜儅僂僗偺憖嶌偱暘巕儅僗僋傪梕堈偵嶌惉偡傞偙偲偑偱偒傞丅(恾2嶲徠)BLA偺暘巕儅僗僋偼丄3侌暘夝擻偺MIR儅僢僾偐傜嶌傜傟偨丅
丂堦曽丄NCS僷儔儊乕僞乕偼丄傑偢廳尨巕僒僀僩偺埵抲偐傜峳偔媮傔丄偙傟傪丄5侌暘夝擻偺MIR儅僢僾偵偍偄偰NCS偺娭學偵偁傞暘巕娫偺揹巕枾搙憡娭偑嵟戝偲側傞傛偆偵丄幚嬻娫僒乕僠傪巊偭偰惛枾壔偟偨丅偝傜偵丄DM傪巊偭偰幚嵺偵averaging傪帠慜偵壗夞傕峴偄側偑傜惛枾壔偟丄偦偺寢壥丄廂懇偟偨抣偑惓妋側NCS僷儔儊乕僞乕偱偁傞偲偟偰丄DM偺杮斣偱巊梡偟偨丅
丂DM偵傛傞幚嵺偺density modification偱偼丄NCS averaging 偵 solvent flattening 偲 histogram matching傪慻傒崌傢偣偰峴偄丄100僒僀僋儖偺偆偪偵暘夝擻傪5偐傜3侌偵忋偘偨丅偙偆偟偰摼傜傟偨3侌暘夝擻偺揹巕枾搙恾偼丄惛枾壔屻偵昤偐傟偨揹巕枾搙恾偺傛偆偵偒傟偄偱偁偭偨丅(恾1嶲徠)偦偙偱丄偙偺BLA偺揹巕枾搙暘晍傪儌僨儖暘巕偲偡傞暘巕抲姺朄偵傛傝BAA偺峔憿夝愅傪帋傒偨丅
丂DM屻偺揹巕枾搙暘晍傪巊偭偨暘巕抲姺朄偵傛傞BAA偺夝愅DM屻偺3侌暘夝擻偺揹巕枾搙暘晍傪巊偭偨BAA偺暘巕抲姺朄偵傛傞夝愅偼丄師偺庤弴偱峴傢傟偨丅
BAA偲BLA偺暘巕儌僨儖偺慻棫丒惛枾壔
丂BAA偼暘巕抲姺朄偵傛傝丄BLA偼丄MIR朄 + density modification l偙傛傝嫟偵椙幙偺揹巕枾搙恾偑摼傜傟偨偨傔丄暘巕儌僨儖偺峔抸偼弴挷偵恑傫偩丅偳偪傜偺応崌傕嵟弶偺暘巕儌僨儖偺峔抸偼丄3侌暘夝擻偺揹巕枾搙恾傪梡偄偰峴偄丄幚嵺偵慻傒棫偰 偨偺偼旕懳徧扨埵拞偺偳偪傜偐1暘巕偺傒偱偁傞丅偙傟傪NCS僷儔儊乕僞乕偱曄姺偡傞偙偲偵傛傝丄傕偆1偮偺暘巕偺儌僨儖傪摼偨丅暘巕儌僨儖偺峔抸丒廋惓偵偼丄僾儘僌儔儉TURBO-FRODO偑梡偄傜傟偨丅
丂暘巕儌僨儖偺惛枾壔偼丄僾儘僌儔儉X-PLOR傪梡偄偰 simulated annealing 偺庤朄偱峴傢傟偨丅暘夝擻偼丄嵟弶偵慻傑傟偨暘巕儌僨儖傪惛枾壔偡傞偲偒偺傒3侌偲偟丄偦傟埲崀偺僗僥乕僕偱偼丄夞愜僨乕僞偺摼傜傟偰偄傞忋尷丄BAA偼2.3侌丄BLA偼2.2侌偱惛枾壔偑恑傔傜傟偨丅2Fo-FC僼乕儕僄恾偑晄柧椖側晹暘偵偮偄偰偼omit map傪昤偒暘巕儌僨儖偺嵞峔抸傪峴偭偨丅悈暘巕偺慻傒擖傟偼丄寢徎妛揑R抣偑20%傪愗偭偰偐傜峴傢傟偨丅嵟廔揑側R抣偼丄BAA偑丄10-2.2侌暘夝擻偺斀幩偵懳偟偰15.7%丄BLA偑丄10-2.3侌偱14.8%偱偁偭偨丅
丂BAA偱偼丄慡483巆婎偵偮偄偰峔憿傪柧傜偐偵偡傞偙偲偑偱偒丄傑偨丄峺慺偺峔憿曐帩偵廳梫側僇儖僔僂儉僀僆儞偺寢崌僒僀僩傕3僇強摨掕偡傞偙偲偑偱偒偨丅偦偺堦曽偱丄BLA偱偼丄億儕儁僾僠僪嵔偺N媦傃C枛抂偺1乣2巆婎偲僇儖僔僂儉僀僆儞偺寢崌偵娭梌偡傞巆婎斣崋180乣190晅嬤偺儖乕僾偺峔憿偑丄僨傿僗僆乕僟乕偺偨傔揹巕枾搙恾忋偵尰傟偰偄側偐偭偨丅傑偨丄僇儖僔僂儉僀僆儞傕寢崌偟偰偄側偐偭偨丅兛亅傾儈儔乕僛偱偼丄僇儖僔僂儉僀僆儞偑丄峺慺妶惈偵昁恵偱偁傞偙偲傪峫偊傞偲丄崱夞摼傜傟偨BLA偺峔憿偼丄妶惈峺慺偺偦傟偲偼庒姳堎側傞傕偺偲峫偊傜傟傞丅側偍丄Huber 傜偑偍偙側偭偨暿偺徎宯偺BLA偺寢徎夝愅1)偱傕丄寢徎壔偺嵺偵EDTA傪巊偭偰偍傝丄椉枛抂偺1乣2巆婎丄180乣190晅嬤偺儖乕僾偺峔憿媦傃僇儖僔僂儉僀僆儞偑尒偊偰偄側偄丅偙偆峫偊傞偲丄崱夞偺BAAL偺峔憿夝愅偵傛傝丄Bacillus 懏桼棃偺峺慺偵偮偄偰弶傔偰偦偺乽妶惈側乿峔憿偑柧傜偐偵偝傟偨偲傕偄偊傞丅
寢壥媦傃峫嶡
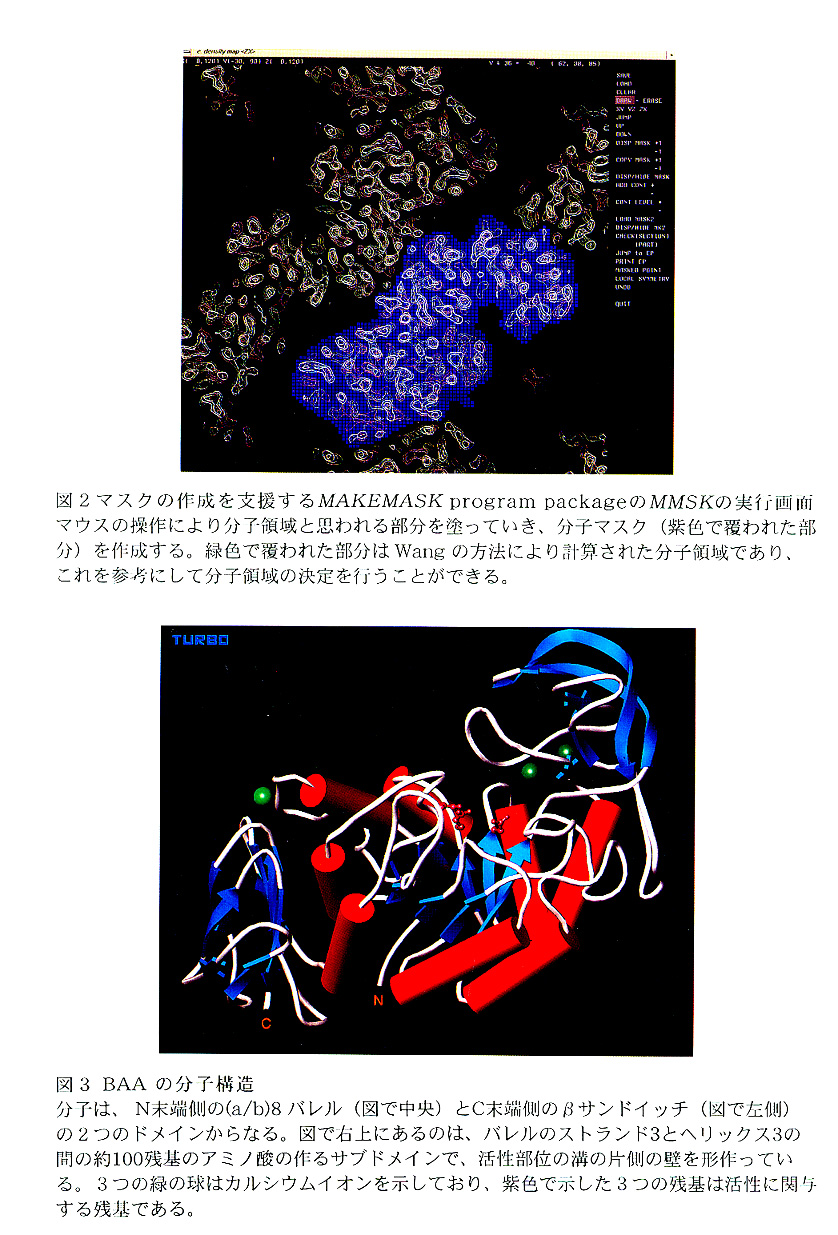
丂暘巕慡懱偺峔憿BAA/BLA偲傕丄暘巕偼丄(兛/兝)8僶儗儖峔憿傪偲傞N枛抂懁僪儊僀儞偲兝僒儞僪僀僢僠峔憿傪偲傞C枛抂懁僪儊僀儞偺2偮偺僪儊僀儞偐傜側傞丅偦偟偰丄妶惈晹埵偼丄(兛/兝)僶儗儖偺兝僗僩儔儞僪偺C枛抂懁偵宍惉偝傟偰偄傞丅(恾3嶲徠)偙偆偟偨暘巕偺婎杮峔憿偼丄墿崓嬠桼棃偺僞僇傾儈儔乕僛3)傗戝敒4)傗僽僞鋁憻桼棃5)偺兛傾儈儔乕僛偲傎傏摨堦偱偁傞偑丄(兛/兝)8僶儗儖偵偍偗傞奺兛僿儕僢僋僗偺挿偝傗憡懳埵抲丄僗僩儔儞僪亅僿僢儕僋僗娫偺儖乕僾偺挿偝傗峔憿偼丄BAA/BLA偲偙傟傜偱戝偒偔堎側偭偰偄傞丅懠偺桼棃偺峺慺偲斾傋偨偲偒偵丄BAA/BLA偺峔憿偵偍偄偰栚傪堷偔晹暘偼丄(兛/兝)僶儗儖偺兝僗僩儔儞僪3偲兛僿儕僢僋僗3偺娫偵懚嵼偡傞栺100巆婎偺傾儈僲巁偐傜側傞峔憿偱偁傞丅偙偺晹暘偼丄妶惈晹埵偺偁傞峚偺曅懁偺暻傪宍惉偡傞偑丄BAA/BLA偱偼偙偺晹暘偑懠偺傾儈儔乕僛偵斾傋偰摿偵戝偒偔敪払偟偰偍傝丄1偮偺僒僽僪儊僀儞傪宍惉偟偰偄傞丅
僇儖僔僂儉寢崌晹埵
丂寢徎夝愅偺寢壥丄BAA偱偼丄寢徎拞偱暘巕1偮摉偨傝3偮偺僇儖僔僂儉僀僆儞偑寢崌偟偰偄傞偙偲偑夝偭偨丅偙偺堦曽偱丄崱夞夝愅偟偨BLA偱偼丄寢徎壔偵EDTA傪梡偄偨偨傔僇儖僔僂儉僀僆儞偺寢崌偼尒傜傟側偐偭偨丅
丂BAA偵偍偄偰丄僇儖僔僂儉僀僆儞偼丄N枛抂懁僪儊僀儞偲C枛抂懁僪儊僀儞偺娫偵1偮偲丄妶惈晹埵偺曅懁偺暻傪宍惉偡傞僒僽僪儊僀儞偺拞偵2偮寢崌偟偰偍傝丄峺慺偺棫懱峔憿曐帩偵巊傢傟偰偄傞丅(恾3嶲徠)偙偺偆偪丄摿偵僒僽僪儊僀儞拞偺2偮偼廳梫偱偁傝丄偙傟傜偺柍偄BlA偱偼丄偙傟傜偵傛傝巟偊傜傟偰偄偨偲峫偊傜傟傞11巆婎傛傝側傞儖乕僾峔憿偑丄揹巕枾搙恾忋偵尰傟偰偍傜偢丄僇儖僔僂儉偑柍偄偙偲偵傛傝僨僀僗僆乕僟乕偵側偭偰偄傞偙偲偑柧傜偐偵側偭偨丅
BAA偲BLA偺峔憿偺斾妑偵傛傞峺慺懴擬壔婡峔偺専摙
丂BAA偲BLA偺帄揔壏搙偺堘偄偼栺25亷偁傞丅傑偨丄90亷偵曐偭偨応崌丄BLA偼BAA偵斾傋栺100攞挿庻柦偱偁傞丅偙偆偟偨椉峺慺偺懴擬惈偺堘偄偼丄棫懱峔憿偵斀塮偝傟偰偄傞傕偺偲峫偊傜傟傞丅偦偙偱丄椉峺慺偺峔憿傪斾妑偡傞偙偲偵傛傝丄BLA偺懴擬惈偺婡峔傪挷傋偨丅椉幰偺庡嵔峔憿偼丄慡C兛尨巕偺r.m.s.曃嵎偑0.50侌偲椙偄堦抳傪帵偡丅(恾4嶲徠)偦偙偱丄椉幰偺娫偱傾儈僲巁抲姺偺婲偙偭偰偄傞暘巕慡懱偺20%偵憡摉偡傞94巆婎偵偮偄偰丄懁嵔偺憡屳嶌梡偺堘偄傪挷傋偨丅偙傟傜偺偆偪偺栺2/3偼丄暘巕昞柺偵埵抲偟丄暘巕撪憡屳嶌梡偵娭梌偟偰偄側偄丅巆傝偺栺1/3(30巆婎)偑丄暘巕撪憡屳嶌梡偵娭梌偟偰偍傝丄廬偭偰BLA偲BAA偱憡屳嶌梡偵壗傜偐偺嵎偑尰傟偰偄傞晹埵偱偁傞丅
丂偙傟傜偺晹埵偱丄悈慺寢崌偺悢偺憹尭傪挷傋偰傒傞偲丄BAA堦>BLA偺曄壔偵傛傝丄懁嵔堦懁嵔娫枖偼懁嵔堦庡嵔娫偺暘巕撪悈慺寢崌偺悢偑丄5憹壛偟2尭彮偟偰偄傞丅偙偺傛偆偵懴擬壔偵敽偆悈慺寢崌偺悢偺憹壛偼偦傟傎偳戝偒偔側偐偭偨丅
丂堦曽丄慳悈惈憡屳嶌梡偵栚傪岦偗傞偲丄BAA堦>BLA偺曄壔偵傛傝丄暘巕撪晹偵偍偄偰恊悈惈傾儈僲巁偑慳悈惈傾儈僲巁偵姺傢偭偨偲偙傠偑柍偔丄BLA偵偍偄偰怴偨偵慳悈惈憡屳嶌梡偑惗偠偨晹埵偼柍偐偭偨丅偨偩偟丄慳悈惈傾儈僲巁偑暿偺慳悈惈傾儈僲巁偵抲偒姺傢傞偙偲偵傛傝丄慳悈惈懁嵔摨巑偺愙怗柺愊偑柧傜偐偵憹壛偟偰偄傞晹埵偑2僇強傎偳尒偮偐偭偰偄傞丅
丂側偍丄BAA->BLA偺曄壔偵傛傝丄怴偨偵僜儖僩僽儕僢僕偼偱偒偰偄側偄丅傑偨丄S-S寢崌偺摫擖傗僾儘儕儞巆婎偺摫擖偵傛傝丄庡嵔偺僐儞僼僆儊乕僔儑儞偵懇敍傪梌偊傞傛偆側曄壔傕婲偙偭偰偄側偐偭偨丅
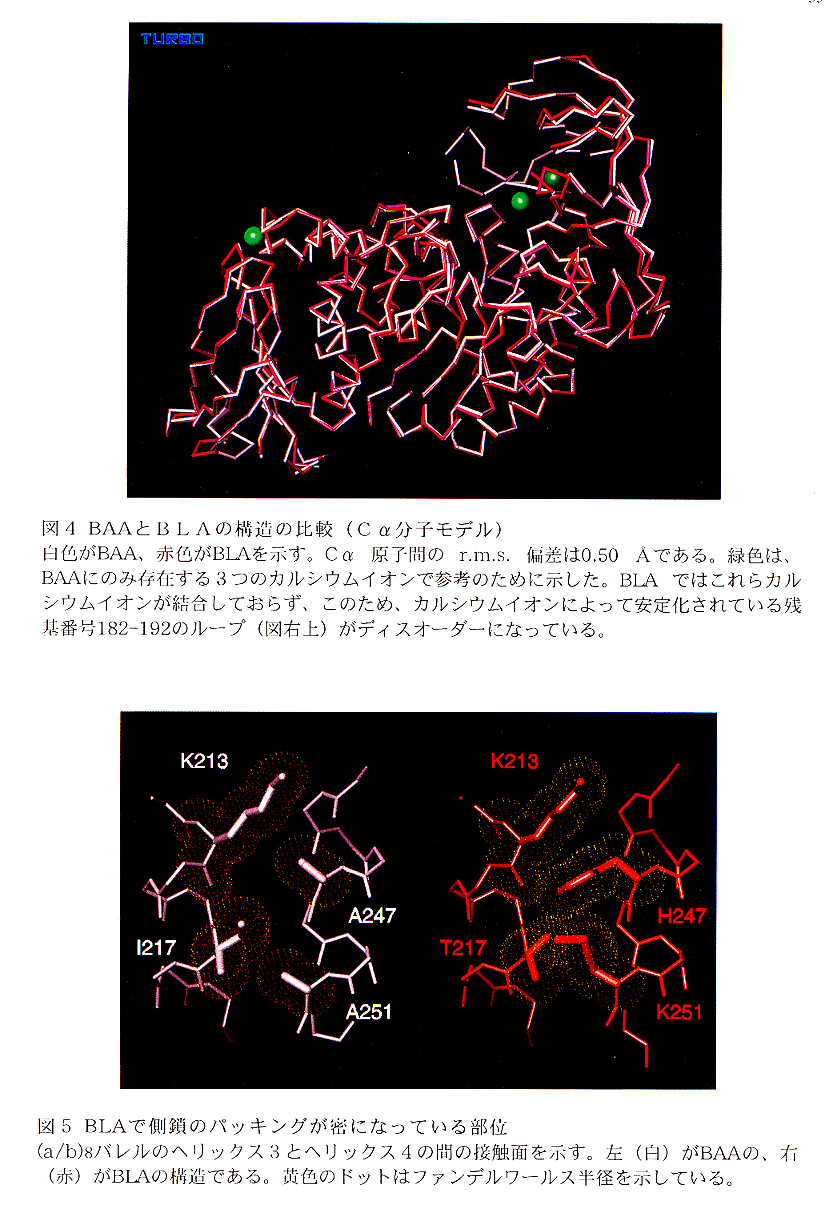
丂BAA偲BLA偺峔憿傪斾妑偟偨偲偒丄椉幰偺傾儈僲巁巆婎偺堘偄偵傛傞嵎偑嵟傕偼偭偒傝偲偟偰偄偨偺偼丄暘巕撪晹偱偺懁嵔偺僷僢僉儞僌偱偁偭偨丅偡側傢偪丄BLA偺傎偆偑丄懁嵔偺僷僢僉儞僌偑岻傒偱偁傝丄暘巕撪晹偵寗娫偑偱偒擄偔側偭偰偄傞丅偙偺嵟傕尠挊側椺偑丄(兛/兝)僶儗儖僪儊僀儞偵偍偄偰3斣栚偲4斣栚偺兛僿儕僢僋僗偑屳偄偵愙偟偰偄傞晹暘偱偁傞丅(恾5嶲徠)偙偺晹暘偱偼丄BAA偺247斣媦傃251斣偺傾儔僯儞偑BLA偱偼丄僸僗僠僕儞偲儕僕儞偵側偭偰偄傞丅偙傟偵傛傝丄悈慺寢崌傗慳悈惈偺憡屳嶌梡偺戝暆側憹壛偼尒傜傟側偄偑丄兛僿儕僢僋僗娫偺寗娫偑杽傑傝丄僿儕僢僋僗摨巑偑僕僢僷乕偱棷傔傜傟偨傛偆偵偟偭偐傝寢傃偮偄偰偄傞丅
丂BLA偺懴擬壔偺儊僇僯僘儉傪暘巕撪憡屳嶌梡偺揰偐傜偙偺傛偆偵挷傋偰傒傞偲丄BAA->BLA偺曄壔偵偲傕側偆悈慺寢崌傗慳悈惈憡屳嶌梡偺憹壛丄媦傃丄暘巕撪晹偱偺懁嵔偺僷僢僉儞僌偺嵟揔壔側偳偺彫偝側曄壔偑愊傒廳側偭偰丄峺慺慡懱偺峔憿傪戝偒偔曄偊傞偙偲側偔擬埨掕惈傪岦忋偝偣偰偄傞條巕偑晜偐傃忋偑偭偰偒偨傛偆偵巚偆丅
偍傢傝偵
丂傛偆傗偔BAA偲BLA偺峔憿夝愅偵堦抜棊偑偮偒丄尰嵼丄崱屻偺尋媶偺揥奐傪峫偊偰偄傞偲偙傠偱偁傞丅岾偄崱夞夝愅傪峴偭偨BAA-BlmA椉寢徎偲傕丄妶惈晹埵偑寢徎奿巕撪偱偺僷僢僉儞僌偵傛傝傆偝偑傟偰偄側偄丅偟偨偑偭偰丄傑偢丄婎幙傾僫儘僌摍傪寢徎拞偵摫擖偟偰偦偺憡屳嶌梡傪挷傋傛偆偲巚偭偰偄傞丅傑偨丄儔僂僄朄偵傛傞峺慺斀墳偺摦揑夝愅偵傕僩儔僀偟偨偄偲巚偭偰偄傞丅
丂嵟屻偲側傝傑偟偨偑丄偙偺尋媶傪巒傔傞偵偁偨傝帋椏偺採嫙媦傃條乆側彆尵傪捀偄偨柤屆壆戝妛擾妛晹偺塋崅廳嶰嫵庼(尰柤梍嫵庼)丄寢徎峔憿夝愅傪恑傔傞偵偁偨傝婱廳側彆尵傪捀偄偨柤屆壆戝妛岺妛晹偺埌揷嬍堦嫵庼(尰柤梍嫵庼)丄嶳崻棽嫵庼丄曻幩岝幚尡傪峴偆偵偁偨傝戝曄偍悽榖偵側傝傑偟偨嶁晹抦暯嫵庼(尰拀攇戝妛)丄拞愳撝巎攷巑(尰杒奀摴戝妛)丄搉绯怣媣攷巑偵姶幱抳偟傑偡丅傑偨丄偙偺尋媶偼丄柤屆壆戝妛岺妛晹偺媽墳梡壔妛戞4島嵗丄尰惗暔婡擻岺妛戞2島嵗傪懖嬈偟偨摗尨塸庽丄埳摗壝復丄惣旜庻峗丄惎堜抦宐巕丄奺柋婔巕丄楅栘弫暯丄崅嫶惤帯丄徖揷壞旤丄奺巵偲偺嫟摨尋媶偱偁傝丄幚嵺偵幚尡傪峴偭偨斵傜斵彈傜偺婱廳側搘椡偺寢壥惉偟悑偘傜傟偨偙偲傪婰偟姶幱傪帵偟偨偄偲巚偄傑偡丅
嶲峫暥專(夝愅偺庤朄丄憰抲丄僾儘僌儔儉偵娭偡傞暥專偼妱垽偝偣偰捀偒傑偟偨)