1. 偼偠傔偵
惗暔偼丄0亷埲壓偺昘偺拞偐傜丄100亷埲忋偺擬悈拞傑偱丄抧媴忋偺條乆側壏 搙娐嫬偺壓偱惗懅偟偰偄傞丅偦傟傜偺惗柦妶摦傪堐帩偡傞偨傔偵丄偦傟偧傟偺抈敀幙 偼惗堢壏搙偺斖埻偱埨掕偵婡擻偱偒傞傛偆偵愝寁偝傟偰偄傞丅抈敀幙偺擬埨掕惈傪寛 掕偯偗傞梫場傪扵偟弌偡偨傔偵丄忢壏嬠偲岲擬嬠偺抈敀幙偺峔憿斾妑偑偙傟傑偱偵昿 斏偵側偝傟偰偒偨丅偦偺寢壥丄偄偔偮偐偺抈敀幙懴擬壔場巕偑採埬偝傟偰偄傞丅偝傜 偵丄晹埵摿堎揑曄堎傗擬椡妛揑偺庤朄傪梡偄傞偙偲偵傛偭偰丄偙傟傜偺懴擬壔場巕偺 岠壥偑丄偄偔偮偐偺抈敀幙偺宯偱妋擣丒掕検壔偝傟偮偮偁傞丅尰嵼偱偼丄偙傟傜偺抦 尒偵婎偯偄偰丄擬埨掕側抈敀幙傪怴偨偵嶌傝弌偡帋傒偑側偝傟偰偄傞丅偟偐偟丄栚揑 捠傝偺抈敀幙傪摼傞偙偲偼崱偩偵擄偟偄偺偑尰忬偱偁傞丅偙偺栤戣傪夝寛偡傞偨傔偵 偼丄惗暔偑峴偭偰偄傞懴擬壔曽朄傪丄傕偆堦搙尒捈偡昁梫偑偁傠偆丅偙傟傑偱偺抈敀 幙懴擬壔偺尋媶偱偼丄傛傝岠壥揑側懴擬壔場巕傪摫偒弌偡偨傔偵丄傛傝崅壏偱惗懅偡 傞惗暔偺抈敀幙傪僞乕僎僢僩偲偟偰偄偨丅偟偐偟丄100亷埲忋偱惗堢偡傞挻岲擬嬠偱 偝偊丄偙傟傑偱偵巜揈偝傟偰偄傞懴擬壔場巕傪棙梡偟偰偄傞偙偲偑丄嵟嬤偺尋媶偵傛 傝柧傜偐偵側偭偰偒偨(Yip傜丄1995;Hennig傜丄1995;Macedo-Ribeiro傜丄1996)丅 偙偺偙偲偼丄怴偨偵懴擬壔場巕傪扵偝側偔偰傕廫暘偵抈敀幙傪懴擬壔偱偒傞偙偲傪帵 嵈偟偰偄傞丅偮傑傝丄偙傟傑偱偲偼堎側傞怴偨側帇揰偐傜偺傾僾儘乕僠偑昁梫偱偁傞丅 惗暔偼條乆側壏搙偱惗懅偟偰偍傝丄偦傟偧傟偺惗堢壏搙偵懳墳偟偨曽朄(埲壓偙傟傪 愴棯偲屇傇)偱敀屓偺抈敀幙傪埨掕壔偟偰偄傞偼偢偱偁傞丅偙偺愴棯傪柧傜偐偵偡傞 偨傔偵偼丄旕忢偵崅壏偱埨掕側抈敀幙偩偗偱側偔丄暿偺懴擬壔儗儀儖傪帩偮抈敀幙偲 挷傋傞昁梫偑偁傞丅偦偙偱丄偙傟傑偱偵偁傑傝拲栚偝傟偰偙側偐偭偨拞摍搙岲擬嬠偺 抈敀幙偵拝栚偟偨丅拞摍搙岲擬嬠偺抈敀幙偺棫懱峔憿傪崅搙岲擬嬠偺傕偺偲斾妑偡傞 偙偲偵傛偭偰丄崅搙偺拞摍搙偺懴擬壔偺堘偄傪柧傜偐偵偡傞偙偲偑偱偒傞丅偙傟l偙傛 偭偰丄抈敀幙傪條乆側壏搙娐嫬傊揔墳偝偣傞偨傔偺丄傛傝岠壥揑側曽朄傪摫偔偙偲傕 偱偒傛偆丅杮尋媶偱偼丄3-僀僜僾儘僺儖儕儞僑巁扙悈慺峺慺(IPMDH)傪梉堦僎僢僩 偲偟丄拞摍搙岲擬嬠 Bacillus coagulans 桼棃偺杮峺慺(BcIPMDH)偺X慄寢徎峔憿夝愅傪峴偭偨丅
2.幚尡丒峔憿寛掕
峺慺偺挷惢丒寢徎壔
杮峺慺偺堚揱巕偼僋儘乕僯儞僌偝傟偰偍傝丄戝挵嬠偱偺敪尰偑壜擻偱偁傞丅 BcIPMDH傪戝検敪尰偟偨嬠傪挻壒攇攋嵱偟丄63亷偺擬張棟偲億儕僄僠儗儞僌儕僐乕 儖暘夋偺屻丄3庬椶偺僇儔儉僋儘儅僩僌儔僼傿乕(Q-sepharose丄Hydroxylapatite丄 Phenyl-5PW)偵傛偭偰寢徎壔梡偺抈敀幙梟塼傪摼偨丅
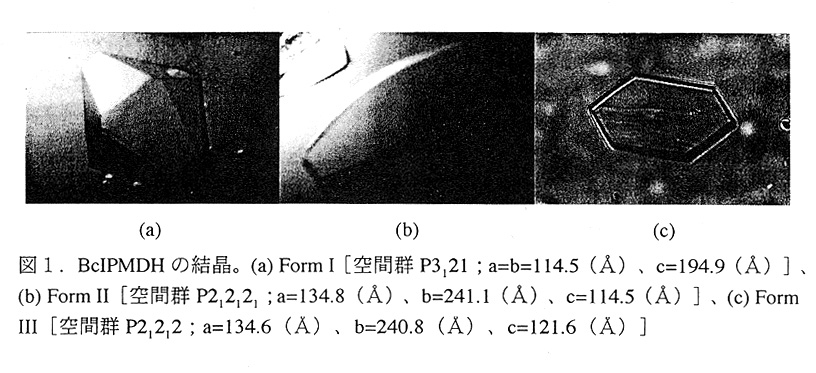
寢徎壔偼僴儞僊儞僌僪儘僢僾忲婥奼嶶朄偱峴偭偨丅棸巁傾儞儌僯僂儉偲億儕 僄僠儗儞僌儕僐乕儖傪捑揵嵻偲偟偰寢徎壔忦審傪専嶕偟偨寢壥丄堎側傞3庬椶偺寢徎 傪摼傞偙偲偵惉岟偟偨(恾侾)丅寢徎偺枾搙應掕偺寢壥丄Form I 偺旕懳徧扨埵偵偼擇 検懱暘巕侾屄(2僒僽儐僯僢僩)傪娷傫偱偄傞偙偲偑傢偐偭偨丅VM(=4.6)偑Matthews 偵傛傝曬崘偝傟偰偄傞抣偵斾傋偰挊偟偔崅偄丅偙偺傛偆偵梟攠娷検偺崅偄寢徎偼 glycolate oxldase(VM=5.0;Lindqvist傜丄1980)傗kallikrein A 偲 trypsininhibitor偺暋崌 懱(VM=4.5;Chen傜丄1983)側偳丄偙傟傑偱偵偄偔偮偐尒偄弌偝傟偰偍傝丄1968擭 偵Matthews偑採埬偟偨VM偺斖埻傪尒捈偡昁梫偑偁傞偩傠偆丅
峔憿寛掕
Form I偑3偮偺寢徎偺拞偱嵟傕崅偄夞愜擻(乣3侌)傪帵偟偨偺偱丄偙偺寢 徎峔憿偺寛掕傪帋傒偨丅X慄夞愜僨乕僞偺廂廤偼丄崅僄僱儖僊乕暔棟妛尋媶強曻幩岝 丄 幚尡巤愝偺嫄戝暘巕梡儚僀僙儞儀儖僌僇儊儔傪梡偄偰峴偭偨丅夞愜僨乕僞偺巜悢偯偗 偍傛傃愊暘偼僾儘僌儔儉WlEIS丄僗働乕儕儞僌偼CCP4偺僾儘僌儔儉(SCALA偍傛傃 AGROVATA)傪梡偄偰偦傟偧傟峴偭偨丅偦偺寢壥丄暘夝擻3侌傑偱偱廂棪85%丄Rmerge偑5.4%偺僨乕僞傪摼傞偙偲偑偱偒偨丅
弶婜埵憡偺悇掕偼丄暘巕抲姺朄偱峴偭偨丅崅搙岲擬嬠 Thermus thermophilus 桼 棃偺杮峺慺(TtIPMDH)偺寢徎峔憿(Imada傜丄1991)偐傜億儕傾儔僯儞偺擇検懱傪 嶌惉偟丄嶲徠暘巕偲偟偰揔梡偟偨丅僾儘僌儔儉AMoRe傪梡偄偰夞揮娭悢偲暲恑娭悢 偺寁嶼傪峴偄丄暘巕偺曽埵偲埵抲傪寛掕偟偨丅偝傜偵丄Romit僾儘僼傽僀儖朄偱夝偺懨 摉惈傪妋擣偟偨丅Romit僾儘僼傽僀儖朄偼丄堦晹偺峔憿傪嶍彍偟偨嶲徠暘巕偐傜寁嶼偝 傟偨R抣(Romit)傪棙梡偟偰暘巕抲姺朄偺夝偺懨摉惈傪専徹偡傞曽朄偱丄挊幰傜偑採 埬丒奐敪偟偨傕偺偱偁傞丅廬棃棙梡偝傟偰偄傞R抣偲偼堎側傝丄偙偺曽朄偱偼嶲徠暘 巕偺峔憿偺乽晹暘揑惓偟偝乿偐傜夝傪敾掕偟偰偄傞偺偱丄1偮偺夝偺拞偵娷傑傟傞晹 暘揑堘偄傪帵偡偙偲偑偱偒偨丅偝傜偵丄杮曽朄偵傛傝摼傜傟偨忣曬偼丄偙偺屻偺峔憿 峔抸傪峴偆忋偱嶲峫偲側偭偨丅
摼傜傟偨弶婜埵憡傪棙梡偟偰揹巕枾搙傪寁嶼偟丄BcIPMDH偺寢徎峔憿傪峔抸 偟偨丅慜弎偺傛偆偵Form I 寢徎偼梟攠娷検偑739%偲旕忢偵懡偔丄傑偨丄旕寢徎妛揑擇 夞懳徧偑偁傞偺偱丄梟攠椞堟暯妸壔丄旕寢徎妛揑擇夞懳徧偵傛傞暯嬒壔側傜傃偵僸僗 僩僌儔儉丒儅僢僠儞僌朄傪棙梡偟偨揹巕枾搙偺夵椙傪僾儘僌儔儉DW傪梡偄偰峴偭偨丅 峔憿偺尨巕僷儔儊乕僞乕偺惛枾壔偵偼丄僾儘僌儔儉X-PLOR傪梡偄偨丅偙偺惛枾壔偺 弶婜抜奒偱偼旕寢徎妛揑懳徧偵傛傞惂栺傪壽偟丄嵟屻偵偼偙偺惂栺傪奜偟偰峴偭偨丅 峔憿偺惛枾壔偲晹暘廋惓傪孞傝曉偟峴偭偨寢壥丄嵟廔揑偵暘夝擻10-3.0侌偱R抣 18.0%丄R free偑24.7%偺寢徎峔憿傪摼傞偙偲偑偱偒偨丅
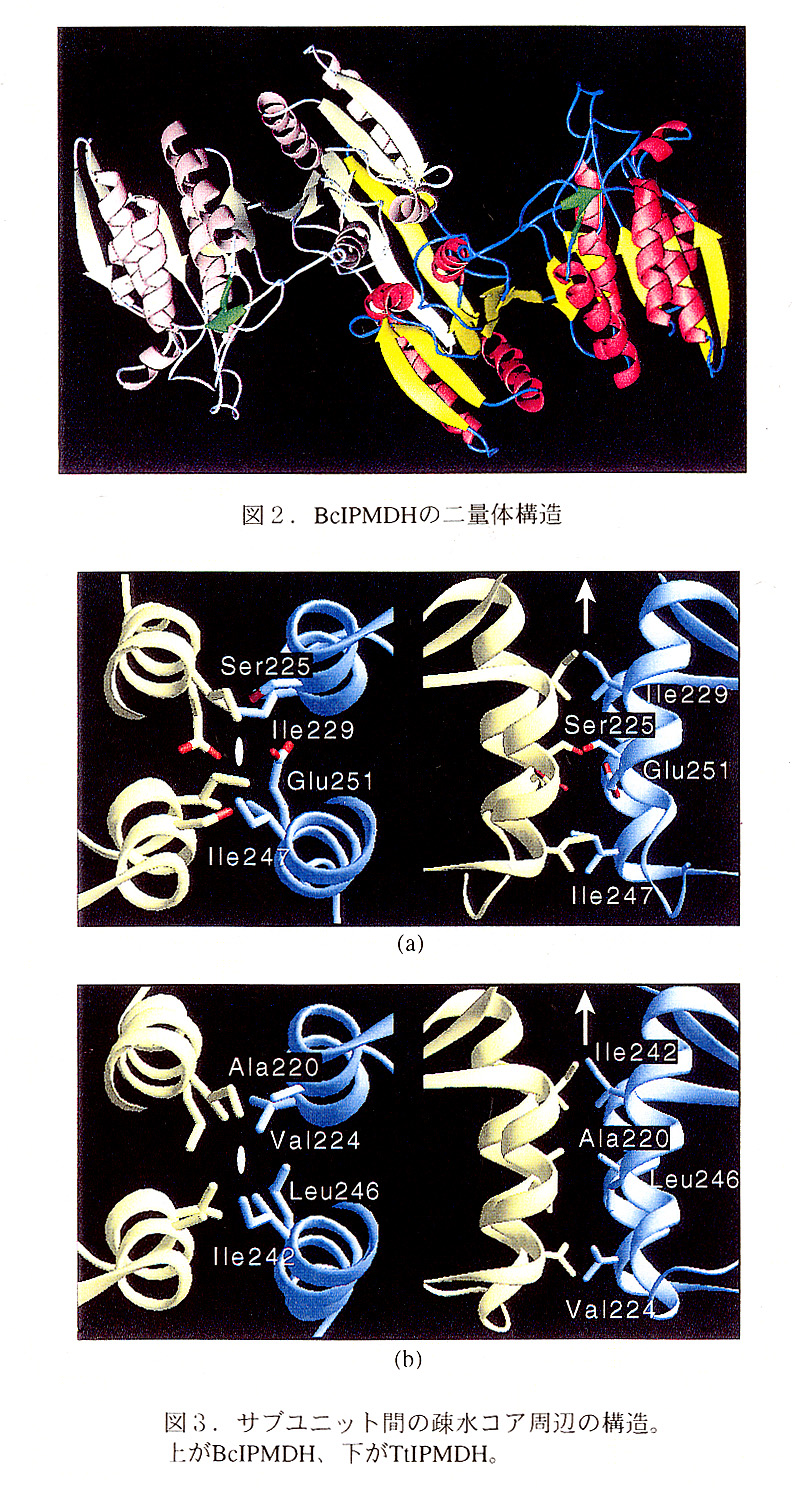
3. 暘巕峔憿
BcIPMDH偼丄暘巕検39808丄傾儈僲巁巆婎悢366偺僒僽儐僯僢僩2偮偐傜側 傞擇検懱暘巕偱偁傞丅慡懱峔憿傪恾2偵帵偡丅奺僒僽儐僯僢僩偼11杮偺兛僿儕僢僋 僗偲14杮偺兝僗僩儔儞僪偐傜側傞open兛/兝峔憿傪偲偭偰偄傞丅偙偺峔憿傪TtIPMDH 偲斾妑偡傞偲丄懳墳偡傞C兛尨巕偳偆偟偺廳偹崌偣偺rms deviation偑1.3侌偲慡懱揑偵 偼椶帡偟偰偄傞丅偟偐偟丄暘巕昞柺偵偁傞偄偔偮偐偺儖乕僾晹暘偱偼丄椉幰偺峔憿偑 挊偟偔堎側偭偰偄偨丅摿偵丄椢怓偱帵偟偰偁傞抁偄兝僔乕僩偼TtIPMDH偵偼側偄擇 師峔憿偱偁傞丅
4. 峫嶡
擬埨掕惈偲懴擬壔場巕
慜愡偱帵偟偨傛偆偵BcIPMDH偲TtIPMDH偺峔憿偵偼戝偒側堘偄偑側偄偵傕 偐偐傢傜偢丄擬埨掕惈偼戝偒偔堎側傞丅偙偺尨場傪扵傞偨傔偵丄傑偢丄偙傟傑偱偵巜 揈偝傟偰偄傞懴擬壔場巕偺柺偐傜2偮偺抈敀幙峔憿傪斾妑偟偨丅
丄 Matthews傜(1987)偼丄Xaa仺Pro枖偼Gly仺Xaa(Xaa偼擟堄偺傾儈僲巁巆婎)
偺抲姺偵傛偭偰曄惈忬懺偺僄儞僩儘僺乕偑尭彮偟丄暘巕慡懱偺懴擬惈傪岦忋偝偣偆傞 偙偲傪巜揈偟偨丅Pro巆婎偼TtIPMDH(25屄)偑BcIPMDH(18屄)傛傝懡偄偺偱丄 偙偺堘偄偑堎側傞懴擬惈偺尨場偺堦偮偲峫偊傜傟傞丅偡側傢偪丄偙偺Pro巆婎悢偺堘 偄偼丄椉幰偺懴擬惈偺娭學偲堦抳偡傞曽岦惈偺曄壔偲偄偊傞丅堦曽丄Gly巆婎悢偵偍 偄偰傕TtIPMDH(36屄)偺曽偑BcIPMDH(31屄)傛傝懡偔側偭偰偄傞偑丄偙偺曄壔 偼曄惈忬懺偺僄儞僩儘僺乕憹戝傪彽偔偺偱丄椉幰偺懴擬惈偺娭學偲偼堦尒柕弬偡傞曽 岦惈偺曄壔偱偁傞丅
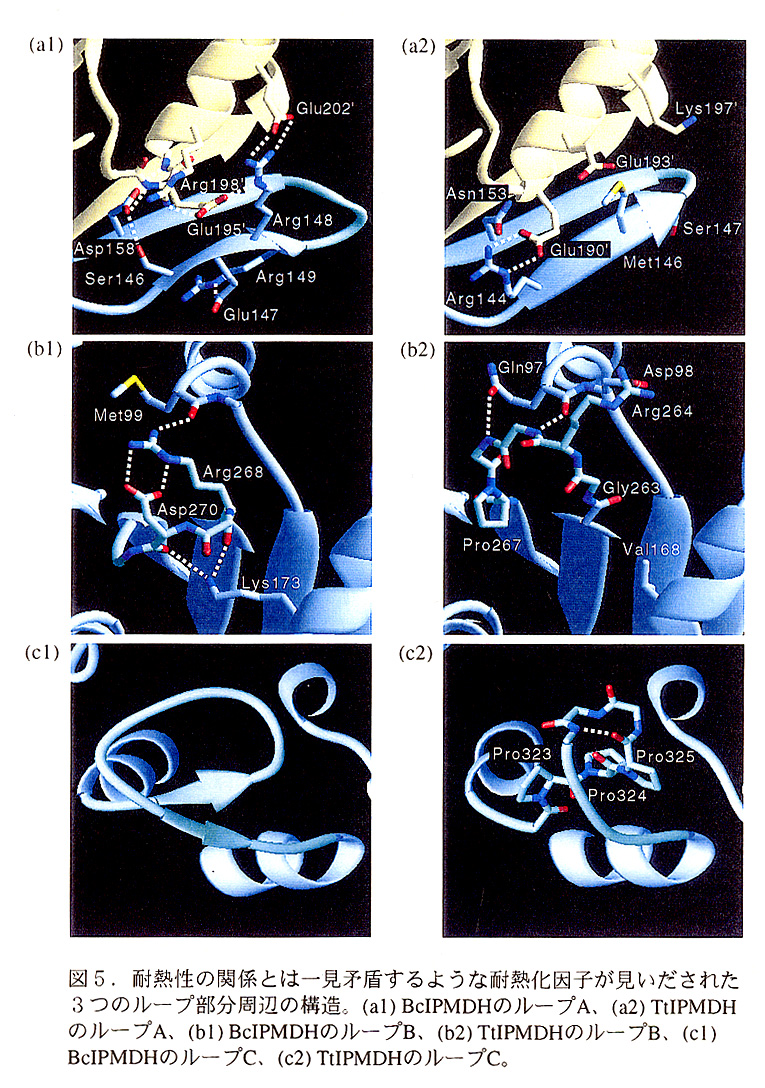
ion-pairdng偲悈慺寢崌偼峔憿偺僄儞僞儖僺乕偵塭嬁傪媦傏偟丄偙傟傜偑懡偄傎 偳暘巕慡懱偺擬埨掕惈偼憹戝偡傞丅抈敀幙撪晹偵杽傕傟偨ion-pair傗悈慺寢崌偺悢偱 偼2偮偺抈敀幙偵戝偒側堘偄偼側偐偭偨偑丄暘巕昞柺偱偼椉幰偲傕偵尠挊側堘偄偑尒 偄弌偝傟偨(昞侾)丅惓晧偺揹壸傪傕偮2偮偺尨巕娫偺惷揹憡屳嶌梡偵傛傝宍惉偝傟 傞ion-pair偼TtIPMDH偺曽偑懡偔丄偙偺曄壔偼2偮偺IPMDH偺懴擬惈偺曽岦偲堦抳 偡傞丅堦曽丄嬌惈尨巕娫偺寢崌嫍棧偲妏搙傪峫椂偟偰悢偊偨悈慺寢崌悢偼BcIPMDH 偺曽偑懡偔丄懴擬惈偺娭學偲偼堦尒柕弬偡傞曽岦惈偺曄壔傪帵偟偰偄偨丅

兛僿儕僣僋僗偺枛抂偼擇師峔憿傪嶌傞悈慺寢崌偑側偄偨傔偵丄偙偺晹暘偵悈 慺寢崌傪摫擖偡傞偙偲偵傛偭偰擇師峔憿傪埨掕壔偟丄傂偄偰偼暘巕慡懱偺懴擬惈傪憹 壛偝偣傞偙偲偑偱偒傞偲曬崘偝傟偰偄傞(Serrano傜丄1989)丅偙傟偼"helix capping" 偲屇偽傟丄Nicholson傜(1988)偼丄尩枾側悈慺寢崌偱偼側偔偰傕丄helix dipole偲憡曗 揑側壸揹傾儈僲巁巆婎傪摫擖偡傞偩偗偱傕懴擬惈偼岦忋偡傞偲庡挘偟偰偄傞丅偙偺 helix capping偼憃曽偺IPMDH偱傕尒傜傟傞偑丄BcIPMDH(16屄)偺曽偑TtIPMDH(12 屄)傛傝埨掕壔偝傟偰偄傞僿儕僢僋僗枛抂偑懡偄丅偙偺曄壔偼丄2偮偺抈敀幙偺懴擬 惈偺娭學偲偼尒偐偗忋柕弬偟偰偄傞丅
抈敀幙撪晹偺慳悈僐傾偵偁傞悈慺寢崌偟偰偄側偄嬌惈傾儈僲巁偼丄暘巕偺擬 埨掕惈傪挊偟偔掅壓偝偣傞(Blaber傜丄1993)丅恾3偵帵偡傛偆偵丄TtIPMDH偺僒僽 儐僯僢僩娫偺愙怗晹埵偵慳悈僐傾偑偁傞丅BcIPMDH偺偙傟偵憡摉偡傞晹暘偱偼丄慳 丄 悈惈傾儈僲巁偑悈慺寢崌偟偰偄側偄Glu巆婎偲Ser巆婎偵抲姺偝傟偰偄傞偺偱丄偙傟 傜偑BcIPMDH偺懴擬惈傪掅壓偝偣傞尨場偺堦偮偱偁傞偲悇應偝傟傞丅偙偺慳悈僐傾 撪偵偁傞傾儈僲巁抲姺偺懴擬惈傊偺塭嬁偼婛偵Imada傜(1991)偑巜揈偟偰偍傝丄Kirino 傜(1994)偵傛傞曄堎懱偺尋媶偐傜傕巟帩偝傟偰偄傞丅
Kellis傜(1988)偼丄暘巕撪晹偵偁傞戝偒側慳悈惈懁嵔傪彫偝側傕偺偵抲姺偡 傞偲暘巕慡懱偺懴擬惈偑尭彮偡傞偙偲偐傜丄偙偺乽寗娫乿偑抈敀幙擬埨掕惈偺堦偮偺 巜昗偲側傝偆傞偙偲傪採埬偟偰偄傞丅偙偺乽寗娫乿偼BcIPMDH(4.4亊102侌3)偺曽 偑TtIPMDH(7.1亊102侌3)傛傝彫偝偔丄偙偺曄壔偼懴擬惈偺娭學偲偼柕弬偡傞曽岦傪帵偟偰偄傞丅
偝傜偵丄傎偐偺懴擬壔場巕偵偮偄偰傕挷傋偨偑丄2偮偺IPMDH偺娫偱尠挊側 堘偄偼尒傜傟側偐偭偨丅
2偮偺懴擬壔愴棯
昞2偼丄BcIPMDH偲TtIPMDH偺峔憿偺堘偄偐傜丄暘巕偺擬埨掕惈偵婏梌偡 傞偲峫偊傜傟傞懴擬壔場巕傪傑偲傔偨傕偺偱偁傞丅偙傟傜偺拞偱丄僾儘儕儞巆婎悢偺 憹壛偲暘巕撪慳悈僐傾偺憹嫮偼丄TtIPMDH偺崅搙側擬埨掕惈偵戝偒偔婑梌偟偰偄傞偲 峫偊傜傟傞丅堦曽丄懴擬惈偺娭學偲偼堦尒柕弬偡傞傛偆側峔憿曄壔(懴擬壔場巕)傕 偄偔偮偐尒偄弌偝傟偨丅偟偐偟丄BcIPMDH偼拞摍搙岲擬嬠偺抈敀幙側偺偱丄拞摍搙 偺擬埨掕壔偺偨傔偵壗傜偐偺懴擬壔場巕傪帩偭偰偄傞偼偢偱偁傞丅偦偙偱丄偙傟傜偺 乽柕弬揰乿偼BcIPMDH偺暿偺懴擬壔曽朄傪帵嵈偟偰偄傞偺偱偼側偄偐偲峫偊丄2偮 偺峔憿傪傕偆堦搙尒捈偟偨丅偦偺寢壥丄暘巕昞柺偵偁傞3偮偺儖乕僾晹暘(儖乕僾A: 150-154丄儖乕僾B:268-272丄儖乕僾C:327-343;偄偢傟傕BcIPMDH偺巆婎斣崋) 廃曈偵偙傟傜偺柕弬揰偑嬌嵼偟偰偄傞偙偲傪尒偄弌偟偨丅偙傟傜偺儖乕僾晹暘偼暘巕 昞柺偵偁傝丄2偮偺抈敀幙偱峔憿偑嬊晹揑偵戝偒偔堎側偭偰偄偨丅偝傜偵丄峺慺偺妶 惈晹埵偐傜棧傟偰偄傞偺偱丄偙傟傜偺嬊晹揑側峔憿曄壔偑峺慺偺婡擻偵偼戝偒偔塭嬁 偟側偄偲峫偊傜傟傞丅埲壓偵丄惗堢壏搙偺堎側傞2偮偺岲擬嬠偑IPMDH傪懴擬壔偡 傞偨傔偵丄偳偺傛偆偵懴擬壔場巕偺巊偄暘偗偰偄傞偺偐傪媍榑偡傞丅

偲偙傠偱丄BcIPMDH偺拞摍搙偺擬埨掕惈傪媍榑偡傞偨傔偵偼丄BcIPMDH傛 傝偝傜偵懴擬惈偺掅偄峺慺傪斾妑偵壛偊傞昁梫偑偁傞丅偦偙偱丄偙偙偐傜偼丄忢壏嬠 Bacillus subtilis 偺IPMDH傪斾妑偵壛偊傞丅偙偺暘巕偺棫懱峔憿偼崱偩偵夝偐傟偰偄側 偄偑丄BcIPMDH偲偺堦師峔憿忋偺憡摨惈偑崅偄(恾4)偺偱丄BsIPMDH偺堦師峔憿 偲BcIPMDH偺嶰師峔憿偐傜BsIPMDH偺棫懱峔憿傪悇應偡傞偙偲偑壜擻偱偁傞偲峫 偊偨丅

BcIPMDH偺儖乕僾B偼5巆婎偐傜側傝丄2偮偺兝僗僩儔儞僪傪宷偖埵抲偵偁 傞丅愭偵巜揈偟偨柕弬揰偲偟偰丄BcIPMDH偺偙偺晹暘偵偼TtIPMDH傛傝懡偔偺悈慺 寢崌偑偁傞(恾5-b1)丅傑偨丄悈慺寢崌枖偼墫嫶傪偟偰偄傞3偮偺巆婎(Lys172丄Arg268丄 Asp270)偼慡偰BsIPMDH偱偼曐懚偝傟偰偄側偄偨傔丄BsIPMDH偵恾5-b1偺傛偆側 憡屳嶌梡偼晄壜擻偱偁傞丅偟偨偑偭偰丄摨堦峔憿傪壖掕偟偰悈慺寢崌傪僇僂儞僩偡傟 偽丄BcIPMDH偺曽偑懡偔側傞偺偱丄暘巕昞柺偵偁傞偙傟傜偺憡屳嶌梡偑BcIPMDH偺 拞摍搙偺擬埨掕惈偵婏梌偟偰偄傞偲悇掕偝傟傞丅堦斒偵丄暘巕昞柺偺悈慺寢崌傗墫嫶 偼擬埨掕惈傊偺岠壥偑彫偝偄偲偝傟偰偄傞偑(Matthews丄1991)丄媡偵丄傢偢偐偵埨 掕惈傪岦忋偝偣傞拞摍搙偺擬埨掕壔偺偨傔偺懴擬壔場巕偱偁傞偲偄偆偙偲傕偱偒傛 偆丅堦曽丄偝傜偵埨掕側TtIPMDH偺偙偺晹暘(恾5-bZ)偵偼暿偺懴擬壔場巕偑偁傞丅 傑偢丄偙偺儖乕僾晹暘偺傾儈僲巁巆婎偺挿偝偑侾巆婎暘抁偔側偭偰偄傞丅儖乕僾晹暘 偺傾儈僲巁巆婎偑抁偔側傞偲曄惈忬懺偺僄儞僩儘僺乕偑彫偝偔側傞偺偱丄擬埨掕惈偵 偲偭偰偼桳棙側曄壔偱偁傞丅傑偨丄挿偔偰廮擃側儖乕僾偼抈敀幙曄惈偵偍偄偰嵟弶偵 峔憿偑曄壔偡傞偲偙傠側偺偱丄抁偄儖乕僾偺傎偆偑曄惈偺奐巒傪杊偖岠壥偑偁傞偲偄 偆曬崘傕偁傞(Russell傜丄1994;Macedo-Rmbeiro傜丄1996)丅傕偆堦偮偺TtIPMDH偺 懴擬壔場巕偲偟偰丄懠偺IPMDH偵偼側偄Pro巆婎偑兝僗僩儔儞僪偺晅偗崻偺晹暘偵 摫擖偝傟偰偄傞丅偙偺曄壔偼丄愭傎偳弎傋偨傛偆偵丄儖乕僾偺廮擃惈傪惂栺偡傞傕偺 偱丄TtIPMDH偺崅搙側懴擬惈偵戝偒偔峷專偡傞偲峫偊傜傟傞丅偟偨偑偭偰丄BcIPMDH 偺儖乕僾B偼丄TtIPMDH偱偼側偖側偭偰偄偨儖乕僾偺挿偝偲廮擃惈傪帩偭偰偍傝丄 拞摍搙偺擬埨掕壔偺偨傔偵悈慺寢崌傪棙梡偟偰偄傞偲夝庍偱偒傞丅偲偙傠偱丄 TtIPMDH偱偼儖乕僾偑抁偔側偭偨偙偲傗丄怴偨側Pro巆婎偺摫擖偵傛傝丄263斣偺庡 嵔偑Gly摿桳偺擇柺妏[(冇丆冋)=(176,-141)]傪偲傜偞傞傪偊側偔側傞丅偟偐偟丄偙偙偼 Gly傊抲姺偝傟偰偍傝丄婲偙傝偆傞棫懱忈奞傪夞旔偟偰偄傞偙偲偑傢偐偭偨丅
儖乕僾A偼丄僒僽儐僯僢僩娫偱兝僔乕僩傪宍惉偡傞偨傔偵昁梫側2杮偺兝僗 僩儔儞僪傪愙懕偟偰偄傞丅偙偺儖乕僾A廃曈晹暘偱傕丄儖乕僾B廃曈偱尒傜傟偨傛偆 側懴擬壔場巕偺巊偄暘偗傪尒傞偙偲偑偱偒傞丅傑偢丄儖乕僾B偲摨條偵丄偙偙偱傕 BcIPMDH偺傎偆偑TtIPMDH傛傝悈慺寢崌偑懡偔側偭偰偄傞(恾5-al)丅BcIPMDH 偺儖乕僾A廃曈偱悈慺寢崌傑偨偼墫嫶傪宍惉偟偰偄傞巆婎偺拞偱丄Glu147偲Arg149 偼BsIPMDH偱偼曐懚偝傟偰偄側偄丅偟偨偑偭偰丄2偮偺巆婎偺娫偺墫嫶偼BsIPMDH 偵偼晄壜擻偱偁傝丄BcIPMDH偺拞摍搙偺擬埨掕惈偵岠壥偑偁傞偲悇應偝傟傞丅堦曽丄 TtIPMDH偱偼丄儖乕僾B偲摨條偵丄悈慺寢崌偑彮側偔側偭偨偐傢傝偵儖乕僾偺抁弅 偲偄偆愴棯傪揔梡偟偰偄傞(恾5-a2)丅偙偺儖乕僾偺抁弅偼BsIPMDH偐傜BcIPMDH 傊偺曄壔偱傕晹暘揑偵尒傞偙偲偑偱偒傞丅傑偨丄BcIPMDH偺儖乕僾A偵偼丄Pro偑1 偮摫擖偝傟偰偍傝丄偝傜偵丄庡嵔偺棫懱忈奞傪夞旔偡傞偨傔偺Gly(151斣)偑偁傞丅 偟偨偑偭偰丄拞摍搙岲擬嬠偺IPMDH偺儖乕僾A晅嬤偱偼丄悈慺寢崌偵傛傞峔憿偺曗 嫮偺傎偐偵丄崅搙岲擬嬠偺IPMDH偺儖乕僾B偱尒傜傟偨儖乕僾偺抁弅偲僾儘儕儞偺 摫擖偵傛傞埨掕壔曽朄偑晹暘揑偵棙梡偝傟偰偄傞丅堦曽丄TtIPMDH偱偼丄庡偵儖乕僾 偺挿偝傪抁弅偡傞偙偲偱懳張偟偰偄傞丅
BcIPMDH偺儖乕僾C偼丄TtIPMDH偵偼側偄抁偄兝僔乕僩偺偁傞晹暘偱偁傞 (恾5-cl)丅忋偵弎傋偨2偮偺儖乕僾廃曈偲摨條偵丄偙偺擇師峔憿偺偨傔偵拞摍搙偵 埨掕側BcIPMDH偺悈慺寢崌偑懡偔側偭偰偄傞丅BsIPMDH偺偙偺晹暘偑摨條偺擇師峔 憿傪宍惉偟偰偄傞偐偳偆偐偼傢偐傜側偄丅偟偐偟丄偙偺晹暘偺堦師峔憿偺椶帡惈偑嬌 抂偵掅偄偺偱丄BsIPMDH偑摨條偺擇師峔憿傪帩偮偳偆偐傪崱屻偺夝愅偱柧傜偐偵偡 傞昁梫偑偁傠偆丅堦曽丄崅搙偵埨掕側TtIPMDH偱偼丄5巆婎暘偺儖乕僾偺抁弅偲丄 3偮偺楢懕偟偨Pro巆婎偺摫擖偵傛偭偰傛傝崅偄埨掕壔傪幚尰偟偰偄傞偲峫偊傜傟傞 (恾5-c2)丅偝傜偵丄TtIPMDH偺偙偺晹暘偱傕丄婲偙傝偆傞棫懱忈奞傪夞旔偡傞偨傔 偺Gly巆婎偑3偮摫擖偝傟偰偄傞丅
偙偙偵帵偟偨BcIPMDH偲BsIPMDH偺3偮偺儖乕僾晹暘偼丄婡擻傪傕偭偨抈 敀幙峔憿傪曐帩偡傞偨傔偵偼乽晄昁梫側儖乕僾偺挿偝乿偲丄Pro巆婎偱懇敍偟偰傕婡 擻偵塭嬁傪偍傛傏偝側偄傛偆側乽夁忚側廮擃惈乿傪帩偭偰偄傞偙偲偑傢偐偭偨丅 BcIPMDH偼丄悈慺寢崌傗墫嫶傪棙梡偟偨峔憿偺曗嫮偵傛傝丄偙傟傜偺晹暘傪埨掕壔 偟偰偄偨丅偙傟偼丄庡偵僄儞僞儖僺乕岠壥偱抈敀幙傪擬埨掕壔偡傞曽朄偱偁傞丅暘巕 昞柺偵偁傞偙傟傜偺憡屳嶌梡偼擬埨掕惈偵戝偒側岠壥偑側偄偲偄傢傟偰偄傞偑 (Matthews丄1991)丄媡偵丄拞摍搙偺擬埨掕壔偺偨傔偵偼揔摉側曽朄偱偁傞偲峫偊傜 傟傞丅堦曽丄崅搙偵埨掕側TtIPMDH偱偼丄乽晄昁梫側儖乕僾偺挿偝乿偲乽夁忚側廮 擃惈乿傪嶍尭偡傞曽朄傪庡偵嵦梡偟偰偄偨丅偙傟傜偼丄偄偢傟傕曄惈忬懺偺僄儞僩儘 僺乕偵塭嬁傪梌偊丄抈敀幙偺懴擬惈傪岦忋偝偣傞偙偲偑偱偒傞丅傑偨丄偙傟傜偺挿偝 傗廮擃惈偼丄嶍傜傟偰傕婡擻偵偼塭嬁偟側偄偲偄偆堄枴偱丄乽峔憿揑側忕挿惈乿偲屇 傇偙偲偑偱偒傞丅偝傜偵丄TtIPMDH偑忕挿惈傪嶍尭偡傞偵偁偨傝丄庡嵔偵棫懱忈奞偑 惗偠傞壜擻惈偺偁傞応崌偵偼丄Gly偺摫擖偱偙傟傪夞旔偟偰偄偨丅
埲忋偺寢壥偐傜丄2庬椶偺嵶嬠偑廃埻偺擬娐嫬偵墳偠偰堎側傞2偮偺抈敀幙 懴擬壔愴棯傪巊偄暘偗偰偄傞偙偲偑傢偐偭偨丅傑偢丄懴擬壔場巕傪摫擖偡傞僞乕僎僢 僩偲側偭偰偄偨偺偼丄忕挿側儖乕僾晹暘偱偁傞丅懴擬壔場巕偺巊偄暘偗曽偼丄懴擬壔 偺儗儀儖偵墳偠偰2捠傝偁偭偨丅拞摍搙偺擬埨掕壔偱偼丄僄儞僞儖僺乕揑愴棯偱偁傞 悈慺寢崌傗墫嫶偵傛傞乽峔憿偺曗嫮乿偑桳岠偱偁傞丅傕偟丄偙偺曗嫮偱抈敀幙傪廫暘 偵埨掕側忬懺偱曐帩偱偒側偔側傞偲丄儖乕僾傪抁弅傗Pro巆婎偺摫擖偲偄偭偨乽峔憿 揑忕挿惈偺嶍尭乿偑昁梫偲側傞丅偙傟偼拞摍搙偺曽朄偲偼堎側傝丄僄儞僩儘僺乕揑愴 棯偱偁傞丅崱屻偼丄杮峺慺傪梡偄偰愴棯偺桳岠惈傪妋擣丒掕検壔偟丄偝傜偵丄懠偺抈 敀幙傊偺墳梡傪帋傒偰偄偔梊掕偱偁傞丅
杮尋媶偼1985擭崰搶嫗岺嬈戝妛棟妛晹偺嶚揷媊晇嫵庼(摉帪)丄栰廆壝柧嫵 庼(尰偄傢偒柧惎戝妛嫵庼)丄娭岥晲巌愭惗(尰偄傢偒柧惎戝妛嫵庼)丄捠嶻徣慇堐 崅暘巕嵽椏尋媶強(尰暔幙岺妛岺嬈媄弍尋媶強)偺忋栰彑旻幒挿偲偺嫟摨尋媶偲偟偰 奐巒偟偨丅尰嵼傕娭岥嫵庼偲偼嫟摨尋媶傪懕偗偰偄傞丅摉弶偼TtIPMDH偺僨乕僞傪 應掕偟偰峔憿夝愅傪恑傔偰偄偨偑丄彅斒偺帠忣偵傛傝夝愅傪抐擮偟偰丄偼偠傔偵弎傋 偨栚揑偵徟揰傪峣偭偰拞摍搙偺擬埨掕惈傪傕偮BcIPMDH偺X慄夝愅偵愗傝懼偊偨丅 BcIPMDH傕斾妑揑抁婜娫偵寢徎壔偡傞偙偲偑偱偒偨偑丄塣埆偔偦傟偼旕懳徧扨埵偵 4暘巕偺僟僀儅乕傪娷傓寢徎偱偁偭偨丅偦偺屻1暘巕偺僟僀儅乕傪娷傓崱夞偺寢徎傪 摼傞偨傔偵悢擭傪旓傗偟偰偟傑偭偨丅偟偐偟丄嬯楯偺峛斻偑偁偭偰旕忢偵嫽枴偁傞寢 壥偑摼傜傟偨偙偲偼朷奜偺婌傃偱偁傞丅X慄夞愜幚尡偱偼崅僄僱儖僊乕暔棟妛尋媶強 偺嶁晹抦暯嫵庼(摉帪)丄拞愳撝巎攷巑(尰杒奀摴戝妛彆嫵庼)丄搉曈怣媣攷巑偵偨 偄傊傫偍悽榖偵側偭偨丅偙偙偵夵傔偰偍楃怽偟忋偘傞師戞偱偁傞丅
嶲峫暥專