遺伝子を分断している、いわゆる、イントロンは、遺伝子操作技術が使えるよう になった1970年代の後半に、真核生物ではグロビン、免疫グロプリン、卵白アルブ ミンなどの遺伝子でまず発見された。現在では、真核生物の多くの遺伝子がイント ロンによって分断ざれてし、ることは周知の事実になっている。最初に発見されたイ ントロンはmRNA型のイントロンであり、mRNAからのイントロンの切り出し(splicing)は、 細胞質に存在するリボ核酸タンパク質(spliceosome)によって行われ る。1つの遺伝子に複数のイントロンが有る場合、切り出すイントロンの数と場所 を変えることで、機能が異なる複数のタンパク質を作ることも可能で、この方法 (alternative splicing)で発生分化の方向を変える現象も多く知られている。
一方、1981年[こなって、酵素の助けを借りないで、RNA分子それ自身でその中にあ るイントロンを切り出す例が、原生生物テトラヒメナのrRNA遺伝子で発見された。 すなわち、self-splicing(セルフスプライシング)するイントロンである。このイ ントロンは、イントロン内の核酸塩基同士が特有の塩基対によって複雑な三次構造 を形成する。ついで、切断される二つの部位が互いに近づいたところで、Mgイ才ン によって触媒されて、イントロン部分が切断除去され、エキソン部分が結合する。 このようにグループ1イントロンは金属酵素の一種である。このイントロンの発見 は、大腸菌のRNase PのRNA部分が真の意味での「酵素」であること(自分白身は変 化しないで、他の分子の変化を触媒すること)の発見と相まって、「RNA酵素」や 「リボザイム」の概念を提出した上[こ、生命系の出現におけるRNAの関与(RNA世界) の存在を示唆するものとなっている。さらに、最近では、RNAウイルス等による病気 の治療にRNA酵素を使う研究が活発になされている。その後の研究、特にミトコンド リアやクロロプラストの研究から、self-splicingするイントロンには、その二次 構造や反応様式の違いから、グループ1イントロンとグループ2イントロンに大別 されている。
グループ1イントロンは、当初の予想以上に生物界に広く分布していることが分 かっている。遺伝子としては、mRNA,tRNA,rRNAと全てのRNAに見つかっている。 また、ミトコンドリアやクロロプラストだけでなく、核ゲノムの遺伝子中にも存在 し、T4ファージや古細菌にまで見つかっている。しかし、今までの所、脊椎動物の 遺伝子では見つかっていない。一方、その分布となると、規則性が見つからず、あ る特定の遺伝子の特定の場所に存在するイントロンも近縁種になくて、遠く離れた 種に存在していたりする場合がある。これに関して、イントロンの「mobility]によ る水平伝達(生物進化に沿った垂直伝達ではなく、生物の種間を越えて水平に伝達 すること)の可能性が論じられている。すくなくとも、酵母のミトコンドリアでは、 特定のイントロンがない個体(I-)とそのイントロンを持つ個体(I+)を接合させて その子孫を調べると、I-の個体にイントロンが移ることが示されている。 また、このイントロンのmobilityに関して、イントロンの中にあるORF(タンパク 質をコードしている部分)の解析が行われ、ある種のORFはsite speciffic DNA endonucleaseであった。このタンパク質酵素がイントロンのないDNA塩基配列を認識 ・切断した後、イントロンのあるDNAを鋳型にした修復反応により、イントロンの mobilityが行われると説明されている(double strand break and repair model)。 私たちは、細胞性粘菌キイロタマホコリ力ビ( Dictyostelim discoideum ,以後 Dicty)のミトコンドリアDNAの構造と機能について、特に、発生・分化とのかかわり や生物進化の観点から研究している。最近、このDNAの全塩基配列を決定し終え、そ の興味深い姿を明らかにした所であるが、ここでは、このDNA中に存在するグループ 1イントロンとそのORFこついて我々が明らかにした点を中心に述べたい。
1)ミトコンドリアのリボソーム大亜粒子RNA(LSU rRNA)遺伝子中のグループ1イ ントロン
T. Cechが1981年[こ、原生生物テトラヒメナのLSU rRNAの前駆体中に存在するイン トロンが、タンバク質分子の助けを借りないでself-splicingしてイントロンを切り 出し、成熟したLSU rRNAになること証明して、RNAが酵素としても働きうることを最 初[こ示した。Dictyの場合も同じよう[こ、この遺伝子の中[こグループ1イントロンが あることが分かった。ただ、このイントロンは、テトラヒメナのイントロンに比ベ てself-splicing活性が低く、生体内ではおそらくタンパク質の助けを借りないとイ ントロンを除去出来ないものと思われる。興味深いことは、Dictyのこのイントロン の挿入箇所は、これまでに調べられたミトコンドリアDNAの中では、単細胞性藻類の Prototheca wickerhamii と緑色藻類の Scenedesmus obliquus と同じであったことで ある。このことは、Dictyが系統的に藻類などに近い側面を持っていることを示唆し ている。これと関連して、上記の2つの生物も遺伝情報は全て普通の暗号を用いて おり、Dictyでも同様に暗号の変化は見つかっていない。

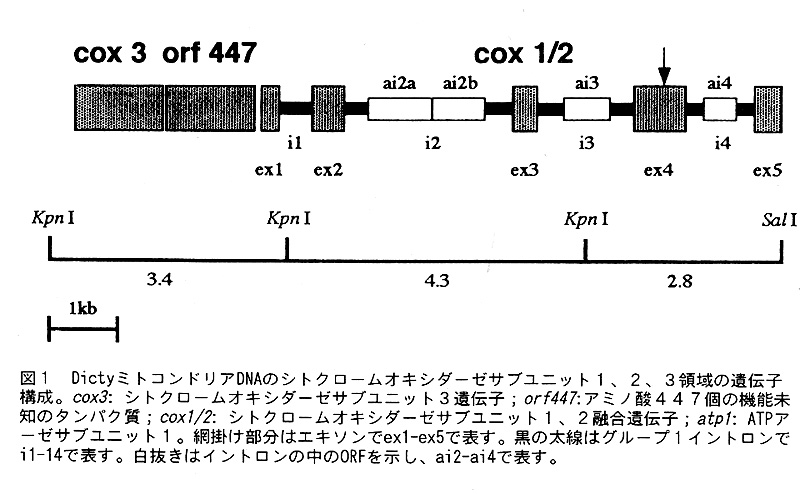
2)シトクローム才キシダーゼ遺伝子中のグループ1イントロン
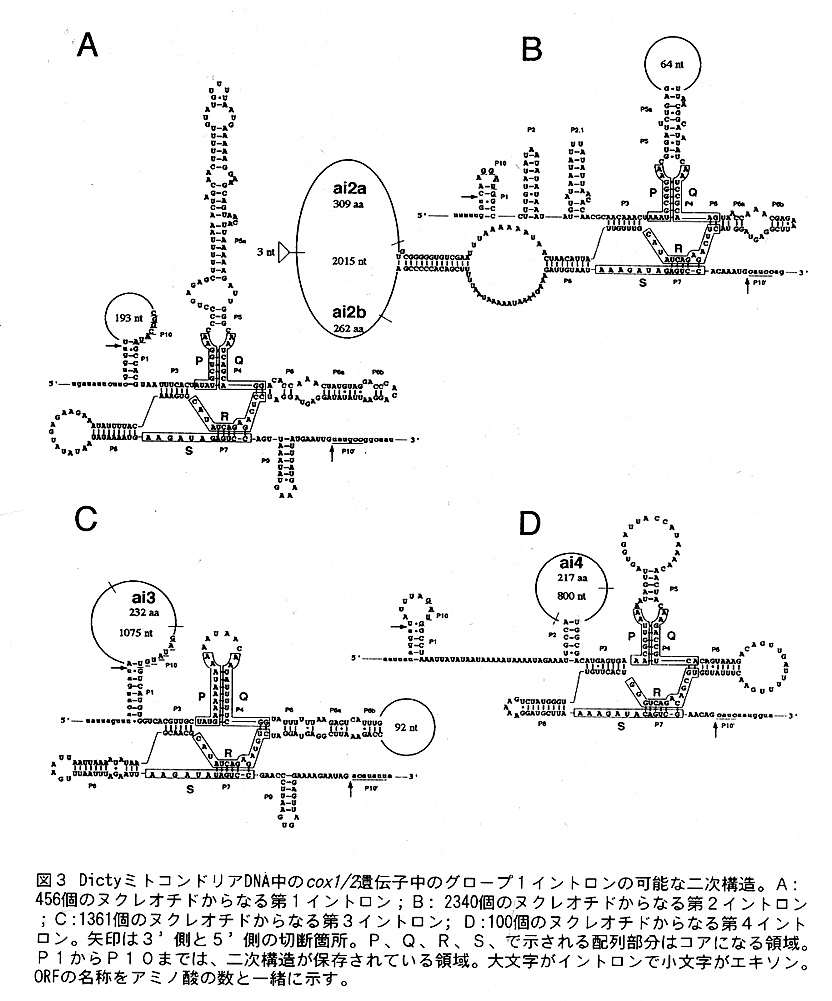
ほとんどの生物のミトコンドリアDNAには電子伝達系の酵素シトクローム才キシダ ーゼサブユニット1、2、3(COX1,COX2,COX3)の遺伝子がコードざれている。こ れらの遺伝子の中には、グループ1イントロンが数多く多く存在しており、例えば、 Podospora anserina lこいたっては、cox1 (遺伝子を表す場合はイタリック体の小文 字で表記する)遺伝子に14個のグループ1イントロンがある。さて、Dictyの場合 は、cox1/2 ( A. castellanii のミトコンドリアDNAと同じよう[こ、cox1 遺伝子とcox2 遺伝子が融合しているので、このように表記する)遺伝子中に4個のグループ1イ ントロンが見つかった。そのうち、3個[こORFが存在していた(図1)。すなわち、 第2イントロンi2にORFのai2aとORFのai2b,第3イントロンi3にORFのai3,第4 イントロンi4にORFのai4である。これらのイントロンのC0X1/2タンパク質の中での 挿入箇所を、他の生物のC0X1とC0X2タンパク質に存在するイントロンの挿入箇所と を比較したのが図2である。P. anserina のai10以外は、同じcoX1遺伝子の相同性の 高い塩基配列の領域の同じ配列の中に挿入されている。図3は、Dictyのこれらのイ ントロンの可能な二次構造を示したものである。これらのイントロン全てがグルー プ1イントロン特徴的な構造を持っている。興味深いのは、第2イントロンi2の同 じループ(L8)のなかに2つのORFが存在することで、この様な例は過去報告されて いない。
3)グループ1イントロンの中のORFの機能
アミノ酸配列の比較から(図4)、イントロン中のORFai2aとORFai2bは両方とも 酵母の cox1 遺伝子中のイントロンのORFあるai4とbi4に似ていることが分かるが、こ の酵母のaf4は、site specificなDNA endonuclease活性を持ち、bi4はグループ1イ ント□ンのself-splicingを助けるRNA maturaseの活性を持つことが分かっている。

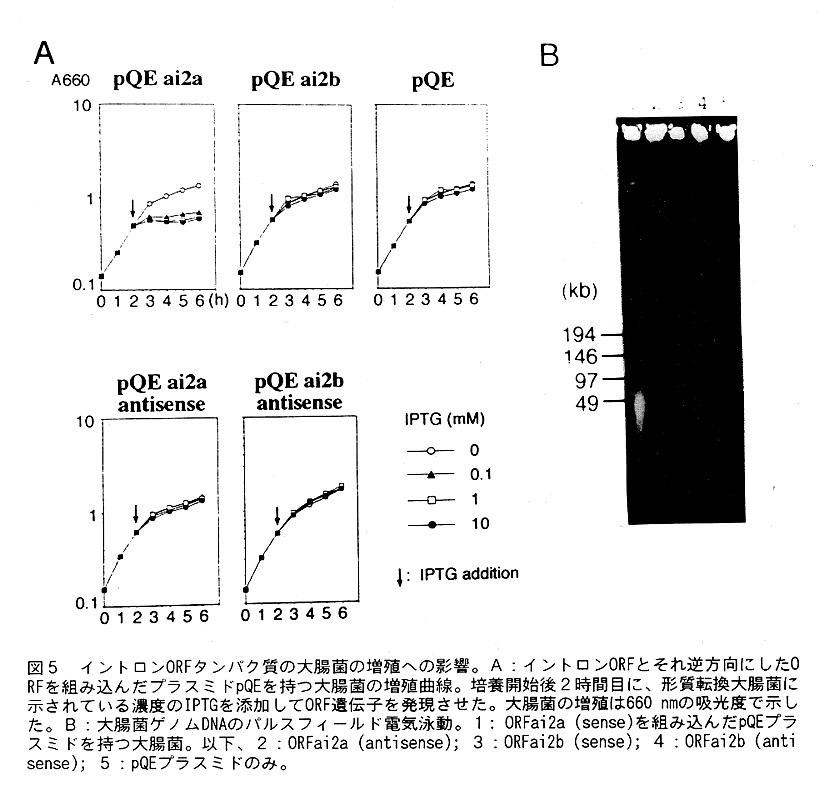
それでは、Dictyのこの2つのORFはどの様な活性・機能を持っているのであろうか。 このことを調べるために、このORFのタンパク質を大腸菌のなかで作らせて精製し、 その活性を調べることにした。pQE60発現ベクターはプロモーターとしてPtac配列を 持っているので、インデユーサーであるラクトースの誘導体IPTGで、そのプロモー タの下流の遺伝子発現を調節出来る。このベクターにフレームが合うように(希望 する正常なタンパク質が作られるように)ORF部分をPCRで調製し、ベクターの特定 の場所に組み込んだ。コントロールとしては、antisenseの配列を組み込んだものを 同様[こして作成した。また、ベクターのみを大腸菌[こいれたものを基本になるコン トロールとして作った。これで、都合5種類の大腸菌の形質転換体を作成して培養 し、培養後2時間目にインデューサーであるIPTGを0.1, 1, 10mM加えた。その増殖 の変化をA660の吸光度で追跡した結果が図5Aである。図から明らかなように、ai2a のORFを正常に組み込んだ場合のみ大腸菌の増殖が阻害されている。そこで、これ らの株からゲノムDNAを調製し、パルスフィールド電気泳動でゲノムDNAの動態を調 べたのが図5Bである。この場合も、ORFai2aを含む大腸菌のみで、約50Kbに断片 化したDNAが検出ざれた。普通、この種のDNA endonucleaseは配列特異的にDNAを切 断する。すなわち、イントロンが挿入されていない(エキソンと次のエキソンが結 合している配列)配列を切断するので、切断されると予想される配列も一緒に組み 込んだ上述のベクターを作成し、同様の実験を行った。大腸菌からベクターDNAを調 製し、ベクターが切断されているかどうかを、まず、EtBrの染色法で調べたが切断 が観察されなかった。より、微量な切断も検出するために、サザンプロット法でも 調べてみたが、それでも、切断していることを示す結果を得ることは出来なかった。 しかし、実際、大腸菌のゲノムDNAが切断されていることは写真から明らかである。 そこで、予想される切断箇所の配列の付近をプライマー伸長法で調べることにした。 もし、ある特定の箇所で切断されていれば、そこでプライマーの伸長が停止しバン ドが形成されることが予想される。この実験をおこないつつあるときに、クラミド モナスのクロロプラストDNA中のSSU rRNA遺伝子中のグループ1イントロン・にコード されている部位特異的なDNA endonucleaseは低いNgイオン濃度では2本鎖を切断でき ず、切断配列の片方の鎖のみを切断することが報告された。また、これと関連して、 クラミドモナスの別の種で、クロロプラストDNA中のLSU rRNA遺伝子中のグループ1 イントロンにコードされているDNA endonucleaseが切断配列の片方の鎖を優先的に 切断することも報告された。
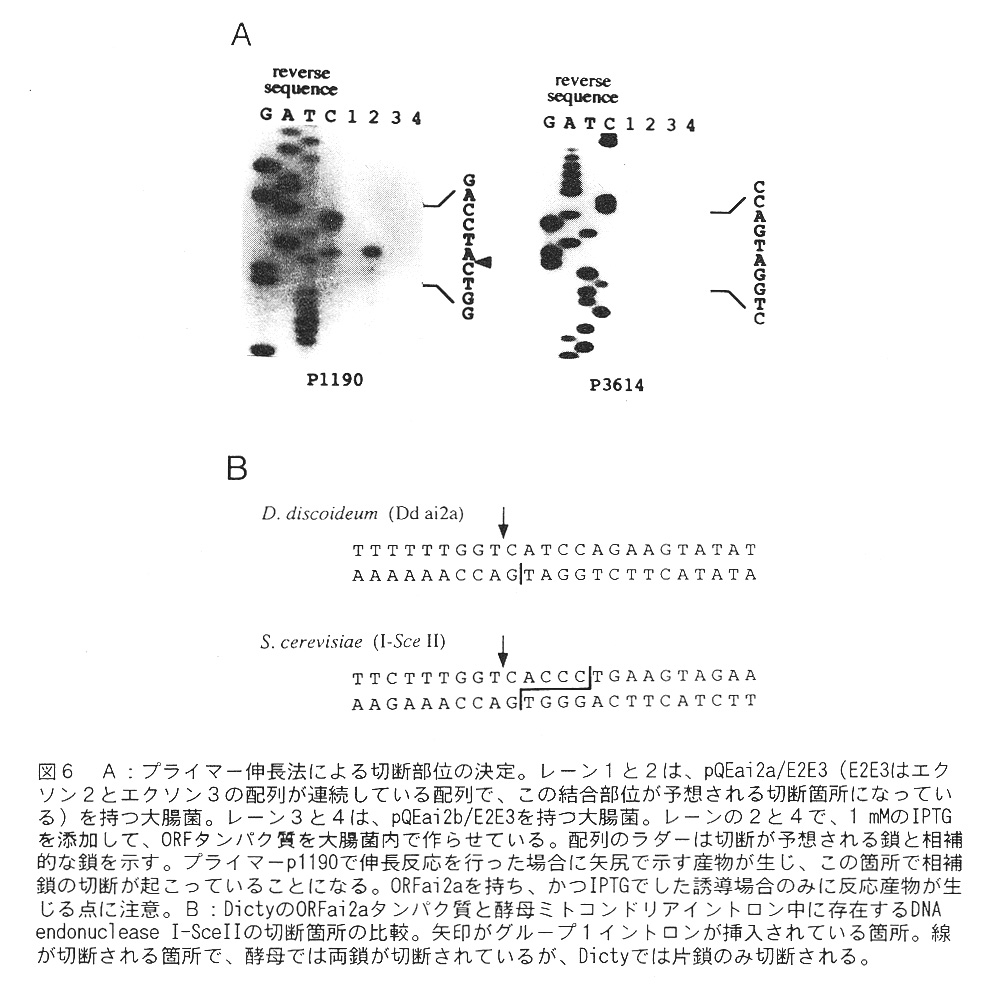
結果は、図6が示す様に、切断箇所の片方のDNA鎖だけを切断することが分かった。 (図6Bの上の鎖だlナが切断ざれている)。興味深いことに、この切断配列は酵母 のsite spesific DNA endonuclease(I-SecII)と似ている上に、切断される箇所が 片方の鎖とは言え、同じであることである(これは、イントロンが cox1 遺伝子の同 じ箇所に挿入されていることから当然であると言えるが)。
一方、Dictyのcox 1/7遺伝子の第2イントロンで、同じループの中にタンデム1こ存 在してし、たORFのaf2bは全く大腸菌の増殖に影響を与えていないので、アミノ酸配列 はa12aに似てはいるが、RNAmaturaseの活性を持つのかもしれないと考えた。この 点を調べるために、精製が簡単なpGEXベクターにORFのai2bを組み込んで、大腸菌で 合成させたこのタンバク質を精製し、第1、第2イントロンのself-splicing反応に 添加して、RNA maturaseの活性が有るかどうかを調べたが、活性を検出することが 出来なかった。これは、すでに、活性を消失してしまったのか、作用するイントロ ンが第1、第2イントロン以外のイントロンであるために活性を示さなかったのか どうかは分からない。また、このai2aとai2bのタンパク質を精製して抗体を作成し、 Dictyのミトコンドリア中にこのタンパク質が存在するかかどうかを調べたが、検出 できなかった。作られる量が極微量のために検出できなかったのか、あるいは、例 えば有性生殖期のような特殊な時期にのみ合成されているのかは、今のところ分か らない。
おわりに
私たちは、DictyのミトコンドリアDNA(55kb)の全塩基配列を決定し解析した。 その結果、そのなかの遺伝子にグループ1イントロンが存在し、イントロンの中に タンパク質をコードしていると考えられる配列があることを見つけた。シトクロー ム才キシダーゼサプユニット1と2の遺伝子の第2イントロンの中に存在するORFの a12aあるいはaf2bは、RNAmaturaseの活性を持つことが予想された。そこで、グル ープ1イントロンと酵素タンパク質との複合体形成や相互作用を調べることを、坂 部プロシェクトの一環として研究することにした。特に、グループ1イントロンは リボザイムの一種である上に、テトラヒメナのグループ1イントロンでは、(1) RNAreplfcaseの活性や、少し改変したこのイントロンでは、(2)endoribonuclease 活性さらには(3)アミノアシルエステル結合の加水分解の触媒活性(この 逆反応でアミノアシルtRNAの合成が可能になる)を持つなど、さらには、グループ 1イントロンはRNA世界で生命系の確立[こ関与したと考えられる興味深いRNA分子で あることなども、この研究に取り組んだ理由である。しかしながら、今までの所、 Dictyのこの2つのORFのタンパク質は、RNA maturase活性を示さない上に、大腸菌 内で多量に作らせることができなかった。今後は、DictyのミトコンドリアDNA中の グループ1イントロンの研究から離れて、このミトコンドリア中に最近見つけた、 129個のヌクレ才チドからなるRNA分子(この分子は、5SrRNAlこ似ているようで似て おらず、リボソームに結合していない)の機能を明らかにするために、このRNAに結 合するタンパク質を精製し始めているところである。
参考文献