偼偠傔偵
C宆娞墛僂僀儖僗(HCV)偺姶愼幰悢偼丄擔杮偩偗偱傕200枩恖埲忋偲悇掕偝傟 偰偄傞丅HCV偼丄姶愼丒敪昦偡傞偲娞峝曄丄娞嵶朎偑傫偲恑峴偟偰偄偔丄擄昦傪堷 偒婲偙偡僂僀儖僗偺傂偲偮偱偁傞丅尰嵼偺偲偙傠C宆娞墛偺帯椕朄偼丄堦晹偵桳岠 惈偺偁傞僀儞僞乕僼僃儘儞椕朄埲奜抦傜傟偰偄側偄丅
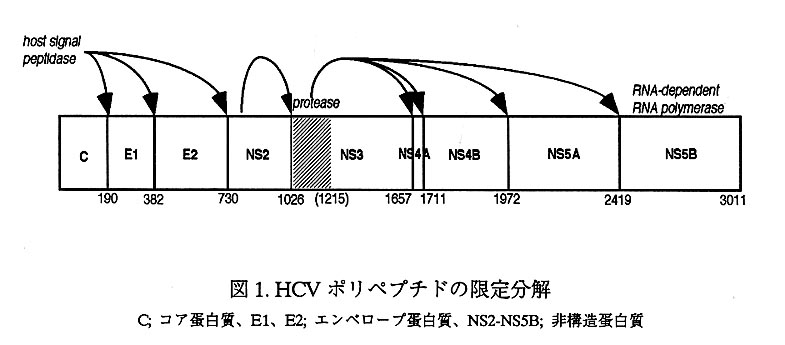
HCV偼墿擬昦僂僀儖僗偵戙昞偝傟傞僼儔價僂僀儖僗偺堦庬偵暘椶偝傟偰偍傝丄栺 9,400墫婎偐傜側傞廫嵔偺堦杮嵔RNA傪僎僲儉偵帩偮丅偙偙偵栺3,010偐傜3,033 傾儈僲巁巆婎傪僐乕僪偡傞挿偄東栿椞堟偑懚嵼偡傞(1)丅東栿偝傟偨億儕儁僾僠僪 偼丄廻庡嵶朎桼棃偺僔僌僫儖儁僾僠僟乕僛偵傛傝尷掕暘夝傪庴偗丄旕峔憿(Non- Structural;NS)抈敀幙偑楢側偭偨NS2-NS3-NS4A-NS4B-NS5A-NS5B偲側傞(恾 1)丅偙偺旕峔憿抈敀幙椞堟偵偼丄偙偺億儕儁僾僠僪傪偝傜偵尷掕暘夝偡傞2庬椶 偺僾儘僥傾乕僛偑僐乕僪偝傟偰偄傞丅傂偲偮偼NS2-3偵傑偨偑偭偨椞堟偵僐乕僪偝 傟偰偄傞嬥懏僾儘僥傾乕僛偱偁傝丄NS2-NS3偺娫傪帺屓徚壔偡傞丅偦偺寢壥丄師偺 僾儘僥傾乕僛(NS3僾儘僥傾乕僛)偑妶惈壔偝傟丄偦偺C枛抂偺NS3-NS4A娫傪僔 僗愗抐偟丄NS4A-NS4B丄NS4B-NS5A丄NS5A-NS5B 娫傪僩儔儞僗愗抐偡傞(2)(恾 1)丅偙偺尷掕暘夝偵傛傝丄椺偊偽RNA埶懚宆RNA億儕儊儔乕僛妶惈傪桳偡傞 NS5B抈敀幙(3)偑妶惈壔偝傟僂僀儖僗偺暋惢偑奐巒偝傟傞丅偙偺傛偆偵丄NS3偺 傕偮僾儘僥傾乕僛妶惈偼丄HCV偺儔僀僼僒僀僋儖偵偍偗傞廳梫側尞偲側偭偰偄傞丅
HIV傗僀儞僼儖僄儞僓僂僀儖僗偺傛偆側懠偺RNA僂僀儖僗偲摨條丄HCV偼暋惢 偺嵺偺撍慠曄堎棪偑崅偔丄儚僋僠儞偺嶌惢偼崲擄偱偁傞丅偙傟偵懳偟偰丄NS3僾儘 僥傾乕僛偺椞堟偼丄HCV庬娫偱偦偺傾儈僲巁攝楍偑摿偵傛偔曐懚偝傟偰偄傞偙偲偐 傜丄偙偺僾儘僥傾乕僛慾奞嵻偼HCV偺帯椕栻偲側傝偆傞壜擻惈偑崅偔丄帯椕栻奐 敪偺廳梫側僞乕僎僢僩偺傂偲偮偲峫偊傜傟偰偄傞丅偡偱偵偄偔偮偐偺尋媶僌儖乕僾 偑丄HCV偺NS3偺僾儘僥傾乕僛椞堟偺棫懱峔憿偺峔憿寛掕偵惉岟偟偰(4丄5)丄 SBDD(Structure Based Drug Design)偵傛傞憂栻偵惗偐偦偆偲偄偆帋傒偑偼偠傑偭 偰偄傞丅
偙偙偱偼丄昅幰傜偑偡偱偵曬崘偟偨慻姺偊懱NS3僾儘僥傾乕僛偺寢徎峔憿(4) 偵婎偯偒丄NS3僾儘僥傾乕僛椞堟偺峔憿丄妶惈晹埵偵偍偗傞婎幙偲偺憡屳嶌梡丄嬥 懏(垷墧)寢崌晹埵偺悇掕丄僾儘僥傾乕僛偺曗場巕偲偄傢傟偰偄傞NS4A偲偺憡屳 嶌梡偵偮偄偰奣愢偡傞丅
1)慻姺偊懱NS3僾儘僥傾乕僛偺惗嶻惛惢偲寢徎峔憿
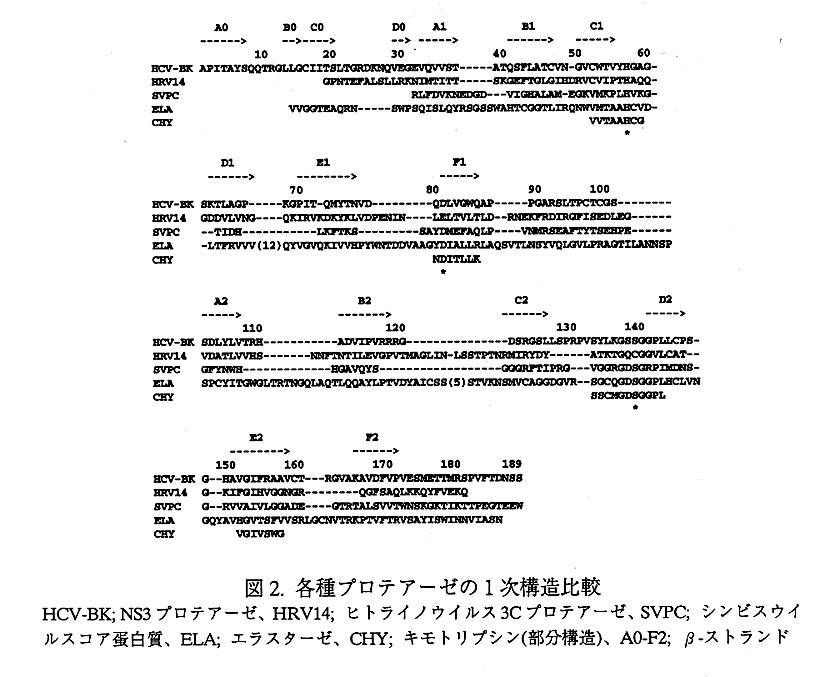
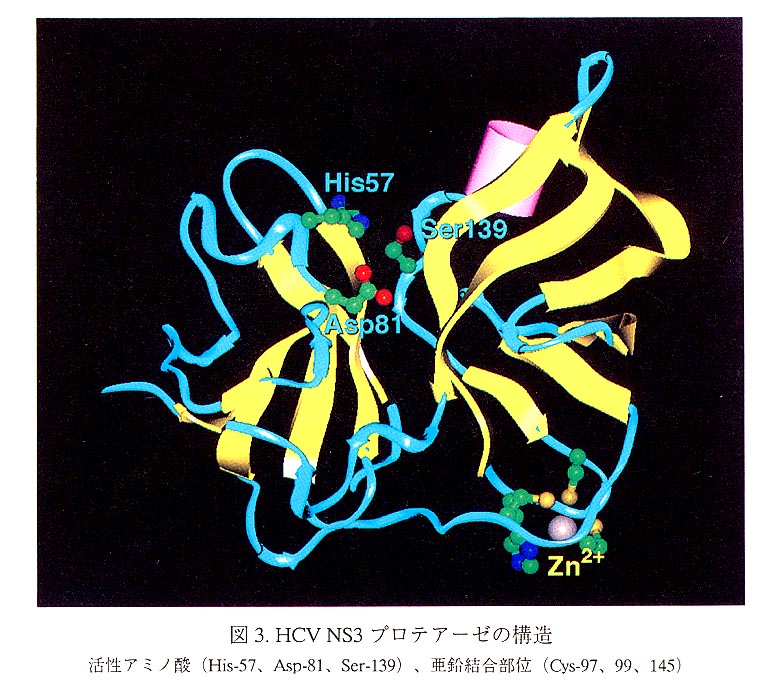
HCV NS3椞堟偼丄631傾儈僲巁偐傜側傝丄偦偺N枛抂偺1/3偺椞堟偵僾儘僥 傾乕僛妶惈偑丄偦偟偰巆傝偺C枛抂2/3偵RNA(DNA)傊儕僇乕僛妶惈偍傛傃 NTP傾乕僛妶惈偑懚嵼偡傞(6)丅傑偨丄N枛抂栺200巆婎拞偺His-57丄Asp-81丄 Ser-139(偦傟偧傟NS3椞堟偺N枛抂偐傜偺斣崋)晅嬤偺傾儈僲巁攝楍偑丄僉儌僩 儕僾僔儞偵戙昞偝傟傞僙儕儞僾儘僥傾乕僛偵昁恵側3偮偺妶惈巆婎(僩儔僀傾乕 僪)晅嬤偺偦傟偲憡摨偱偁傞丅偦偙偱丄NS3僾儘僥傾乕僛偑僙儕儞僾儘僥傾乕僛偱 偁傞偲梊應偝傟丄偙偺3偮偺傾儈僲巁巆婎偑妶惈僩儔僀傾乕僪偱偁傞偙偲偑悇應偝 傟偰偄偨丅偲偙傠偑丄偙偺僩儔僀傾乕僪晅嬤傪彍偔傾儈僲巁攝楍偼懠偺僾儘僥傾乕 僛偲偺憡摨惈偑掅偔丄抈敀幙慡懱偺峔憿傪梊應偡傞偙偲偼崲擄偱偁偭偨(恾2)丅
偦偙偱丄NS3偺偳偺椞堟偑僾儘僥傾乕僛妶惈傪扴偆偺偐丄偁傞偄偼NS3慡椞堟偑 偦偺妶惈偺敪尰偵昁恵偱偁傞偺偐偵偮偄偰偺尋媶偑惙傫偵峴側傢傟偨丅NS3偺N枛 抂偍傛傃C枛抂偐傜僔僗僥儅僥傿僢僋偵抁弅偟偨堚揱巕傪梡偄偨慻姺偊懱抈敀幙偵 傛傞僂僀儖僗偺億儕儁僾僠僪偺僾儘僙僔儞僌傪傒傞偙偲偱丄偦偺僾儘僥傾乕僛妶惈 偵昁梫側椞堟偑専摙偝傟偨丅
昅幰傜偼丄4偮偺戙昞揑側HCV偺H丄BK丄J6丄J8宆堚揱巕傪梡偄丄慻姺偊懱 NS3僾儘僥傾乕僛惗嶻傪帋傒偨丅條乆側慻姺偊懱傪専摙偟偨寢壥丄壓墦栰傜偺僾儘 僥傾乕僛妶惈偺昁恵椞堟偵娭偡傞曬崘(7)偵婎偯偒丄BK宆堚揱巕偐傜1-189傾儈 僲巁椞堟偑僾儘僥傾乕僛妶惈偺偁傞抈敀幙偲偟偰戝挵嬠偱惗嶻偝傟傞帠傪尒弌偟偨丅 偦偟偰丄偦偺崅偄摍揹揰傪峫椂偟偰梲僀僆儞岎姺懱側偳偺僇儔儉僋儘儅僩僌儔僼傿 堦偵傛傝丄僾儘僥傾乕僛妶惈偺偁傞扨堦抈敀幙偲偟偰惛惢偟丄偙偺NS3僾儘僥傾乕 僛偺寢徎壔偲峔憿寛掕傪峴偭偨丅
夝偐傟偨寢徎峔憿偐傜丄NS3僾儘僥傾乕僛偼6偮偺兝僗僩儔儞僪偐傜側傞2偮偺 僶儗儖峔憿偑岦偒崌偭偨丄僉儌僩儕僾僔儞傗僩儕僾僔儞側偳偵尒傜傟傞揟宆揑側僙 儕儞僾儘僥傾乕僛峔憿傪偲偭偰偄傞偙偲偑柧傜偐偵偝傟偨(恾3)丅堦曽丄189傾 儈僲巁偐傜側傞偙偺NS3僾儘僥傾乕僛偱偼丄妶惈晹埵偺峚偵撍偒弌偟偨偦傟偧傟2 杮偺兝僗僩儔儞僪偵傛偭偰宍惉偝傟傞儖乕僾偼丄偍傛偦220傾儈僲巁偐傜側傞僉儌 僩儕僾僔儞傗僩儕僾僔儞偵斾傋偰抁偔丄栺30傾儈僲巁巆婎暘彫偝偄偨傔偵丄僐儞僷 僋僩偱宱嵪揑偵傑偲傑偭偰偄傞(恾2)丅抈敀幙崪奿偺慡懱偺愜傟忯傒偼丄歁擕摦 暔桼棃偺僙儕儞僾儘僥傾乕僛傛傝傕丄僾儘僥傾乕僛偲偟偰僐儞僷僋僩側儔僀僲僂僀 儖僗偺3C僾儘僥傾乕僛傗僔儞價僗僂僀儖僗偺僐傾抈敀幙(僾儘僥傾乕僛)偵嬤偄丅
2)妶惈晹埵偺峔憿
NS3僾儘僥傾乕僛偺寢徎峔憿偱偼丄S1億働僢僩偼懁嵔偺戝偒偄旕嬌惈傾儈僲巁 Phe-154偺朏崄娐偑丄億働僢僩偺拞墰墶偐傜撍偒弌偰偍傝丄偝傜偵Ala-157傗Leu-135偺懁嵔傕傗偼傝墶偐傜怢傃偰偍傝丄P1晹埵偺懁嵔偑寢崌偡傞寠偺戝偒偝偼僉儌 僩儕僾僔儞丄僩儕僾僔儞偺S1億働僢僩傛傝愺偄丅偟偨偑偭偰丄婎幙偲側傞旕峔憿 抈敀幙偺愗抐晹埵偺P1晹埵偵偁傞僩儗僆僯儞偁傞偄偼僔僗僥僀儞巆婎偺彫偝側傾 儈僲巁懁嵔偑偆傑偔擺傑傞戝偒偝偱偁傞丅偡偱偵弎傋偨傛偆偵丄妶惈晹埵傪宍惉偡 傞傆偨偮偺僪儊僀儞娫偵撍偒弌偨儖乕僾偑慡懱偵抁偔側偭偰偄傞偨傔偵丄婎幙偺 P2-P5晹暘偲憡屳嶌梡偡傞抈敀幙昞柺傕僉儌僩儕僾僔儞側偳偲斾傋丄側偩傜偐偱峀 偄丅堦曽丄婎幙偲側傞億儕儁僾僠僪愗抐晹埵偺P6偐傜P2'偵偁偨傞傾儈僲巁攝楍 偼丄BK宆HCV偺応崌丄3-4A丄4A-4B丄4B-5A偦偟偰5A-5B晹埵偱丄偦傟偧傟丄 DLEVVT/ST丄DEMEEC/AS丄DCSTPC/SG丄EDVVCC/SM偱偁傞丅偙偙偱丄 P6埵偺巁惈傾儈僲巁丄P1埵偺僩儗僆僯儞偁傞偄偼僔僗僥僀儞丄偦偟偰P1乫偺僙儕儞 偁傞偄偼傾儔僯儞偑偔傞偙偲傪彍偗偽丄P2-P5晹埵偵傾儈僲巁偺堦師攝楍偵尒傞傋 偒憡摨惈偼側偄丅
偝傜偵S1億働僢僩晅嬤偺峔憿傪懠偺僾儘僥傾乕僛偲斾妑偡傞偲丄NS3僾儘僥傾乕 僛偵偍偄偰偼丄Ala-157偲Val-167偑Sl億働僢僩偺擖岥晅嬤偵偁傝丄億働僢僩偺娫岥 傪婯掕偟偰偄傞丅偙傟傜偵憡摉偡傞傾儈僲巁巆婎偼丄僉儌僩儕僾僔儞偲僩儕僾僔儞 偱嫟偵僌儕僔儞巆婎偲側偭偰偍傝丄婎幙偺P1偵摿堎揑側巆婎偼偦傟偧傟僠儘僔儞丄 僩儕僾僩僼傽儞偦偟偰儕僕儞丄傾儖僊僯儞側偳偱偁傝丄戝偒側懁嵔傪庴偗恖傟傞怺 偄億働僢僩傪帩偮丅偙傟偵懳偟偰丄P1偺婎幙摿堎惈偑傾儔僯儞側偳偺傛偆側彫偝 偄慳悈惈懁嵔傪傕偮傾儈僲巁巆婎偺僄儔僗僞乕僛偱偼丄偙傟傜偺傾儈僲巁巆婎偼偦 傟偧傟僶儕儞偲僩儗僆僯儞偲側偭偰偄傞丅
妶惈僙儕儞(Ser-139)晅嬤偺傾儈僲巁攝楍偼丄NS3僾儘僥傾乕僛偵偍偄偰傕丄 Gly-Ser-Ser-Gly-Gly偲側偭偰偍傝丄僩儕僾僔儞條僾儘僥傾乕僛偵摿挜揑側GXSGG 儌僠乕僼偲側偭偰偄傞(恾2)丅僾儘僥傾乕僛偵傛傞壛悈暘夝偺拞娫懱丄巐柺懱慗 堏忬懺(僆僉僔傾僯僆儞)傪埨掕壔偝偣傞偲偄傢傟偰偄傞135偐傜139偺傾儈僲巁 巆婎偺儁僾僠僪崪奿偐傜側傞儖乕僾傗丄150-158偺傾儈僲巁巆婎偐傜側傞E2偺兝僗 僩儔儞僪偑丄婎幙寢崌晹埵傪宍惉偟偰偄傞丅傑偨丄懠偺僾儘僥傾乕僛偵傒傜傟側偄 C2-D2娫偺儖乕僾偵懚嵼偡傞1僞乕儞暘偺兛傊儕僢僋僗(131-134)偼丄偙偺僾儘僥傾 乕僛偺摿挜偱偁傝丄僆僉僔傾僯僆儞儂乕儖傪宍惉偡傞儖乕僾偺捈慜偵埵抲偡傞丅偙 偺傊儕僢僋僗宍惉偵傛傝丄婎幙寢崌晹埵偑峔憿揑偵埨掕壔偟偰偄傞壜擻惈偑偁傞丅
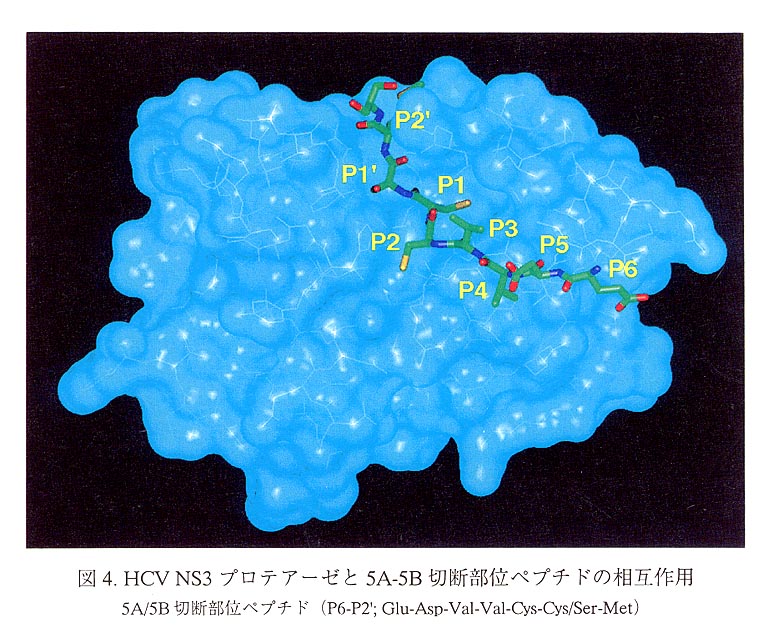
埲忋偵弎傋偨峔憿忋偺摿挜偵婎偯偒丄5A-5B晹埵偺婎幙儁僾僠僪(P6偐傜P21傑 偱)偑NS3僾儘僥傾乕僛偵寢崌偟偨暋崌懱儌僨儖傪帵偡(恾4)丅偙偺儌僨儖傛傟 偽丄S1億働僢僩偵偼P1偺僔僗僥僀儞懁嵔偑擺傑傝丄Phe-154偺揹巕嫙梌惈偱偁傞 儀儞僛儞娐偺兾揹巕塤偲僔僗僥僀儞巆婎偺僠僆乕儖婎偑峀媊偺悈慺寢崌傪宍惉偟偰 偍傝丄P1偺婎幙摿堎惈傪偆傑偔愢柧偱偒傞丅P2-P3偺晹暘偼丄側偩傜偐側NS3僾儘 僥傾乕僛偺妶惈晹埵晅嬤偺偔傏傒偵墶偨傢偭偰偄傞丅偡傋偰偺億儕儁僾僠僪愗抐晹 埵偺P6巆婎偼晧偵壸揹偟偨巁惈傾儈僲巁偲側偭偰偄傞偑丄儌僨儖拞丄P6偺僌儖僞 儈儞巁懁嵔偺僇儖儃僉僔儖婎嬤朤偵偼丄墫婎惈傾儈僲巁巆婎偺Arg-161偲Lys-165 偺惓偵壸揹偟偨懁嵔偑偁傝丄婎幙偺寢崌偵婑梌偟偰偄傞偲峫偊傜傟傞丅
3)垷墧寢崌儌僠乕僼
HCV偺億儕儁僾僠僪偺僾儘僙僔儞僌夁掱傪嵞尰偟偨帋尡娗撪抈敀幙崌惉宯傪梡 偄偨僾儘僥傾乕僛妶惈偼丄悈嬧傗摵側偳偺廳嬥懏僀僆儞偵傛傝梷偊傜傟(8)丄傑偨 垷墧揧壛偵傛傝懀恑偝傟傞丅傑偨丄慻姺偊懱NS3僾儘僥傾乕僛偺妶惈應掕偺嵺丄 EDTA摍偺僉儗乕僩嵻偑妶惈傪慾奞偡傞偙偲傕抦傜傟偰偍傝丄嬥懏僀僆儞偑偙偺 NS3偺僾儘僥傾乕僛妶惈偵怺偔娭梌偟偰偄傞偙偲偑悇應偝傟偨(9)丅
NS3偺僾儘僥傾乕僛椞堟189傾儈僲巁巆婎拞偺攝楍傪H丄BK丄J6丄J8:宆偺娫偱 斾妑偟偰傒傞偲丄BK宆偱7偮偁傞僔僗僥僀儞巆婎偺撪丄Cys-97丄99丄145丄159偑 曐懚偝傟偰偄傞丅晹埵摿堎揑曄堎懱傪梡偄偨妶惈應掕偐傜丄Cys-97丄99丄145偑僾 儘僥傾乕僛妶惈偵昁恵偱偁偭偨丅曐懚偝傟偰偄傞Cys-159傪娷傓4偮偺傾儈僲巁曄 堎懱(C16A丄C47S丄C52L偦偟偰C159S)偼丄栰惗宆偲摨摍偺僾儘僥傾乕僛妶惈 傪桳偟偰偄偨(枹敪昞僨乕僞)丅嵟嬤丄栰惗宆暲傃偵4傾儈僲巁曄堎懱偼丄抈敀幙 暘巕偁偨傝1暘巕摉検暘偺垷墧傪娷桳偟偰偄傞偙偲偑幙検暘愅寁偵傛傞尦慺暘愅偺 寢壥偐傜柧傜偐偲側偭偨(9)丅
NS3僾儘僥傾乕僛偺寢徎峔憿偵偍偄偰丄垷墧寢崌晹埵偺Cys-97丄99丄145偺3偮 偺僔僗僥僀儞巆婎偺棸墿尨巕偲垷墧偺尨巕娫嫍棧偼丄2.0偐傜2.5侌偲側傝丄奺尨巕 偼傎傏惓巐柺懱偺捀揰偵埵抲偟丄偦偺嬤朤偵His-149偑懚嵼偡傞(恾3)丅偙偺垷 墧寢崌晹埵偼丄峺慺妶惈晹埵偐傜墦偔棧傟偰偍傝丄怗攠婡擻偵捈愙娭梌偟偰偄傞偲 尵偆傛傝丄抈敀幙偺峔憿曐帩偵婑梌偟偰偄傞傕偺偲巚傢傟傞丅偙偺qs-97丄99丄145 偲His-149偵戙昞偝傟傞摿挜揑側CXC---CXXXH偁傞偄偼CXH偺僉儗乕僩儌僠乕 僼偼丄懠偵丄HGV丄GB僂僀儖僗丄儔僀僲僂僀儖僗丄億儕僆僂僀儖僗偺僾儘僥傾乕 僛偵傕尒偮偐偭偰偍傝丄怴偟偄僞僀僾偺垷墧寢崌儌僠乕僼偺壜擻惈偑偁傞(9)丅
4)NS4A抈敀幙偺塭嬁
HCV偼丄懠偺僼儔價僂僀儖僗偵偼側偄NS4A偲偄偆54巆婎偐傜側傞彫偝偄抈敀 幙傪僐乕僪偟偰偄傞丅偙偺抈敀幙偼NS3偺僾儘僥傾乕僛妶惈傪忋徃偝偣傞曗場巕偲 偟偰偺婡擻傪帩偮(10)丅摿偵丄NS4A/4B偺娫偺愗抐傪嫮偔懀恑偟丄NS4B15A偺娫 偺愗抐偵偼昁恵偱偁傞偙偲偑抦傜傟偰偄傞(11)丅NS4A偼丄NS3抈敀幙偺N枛抂 晹埵偲寢崌偟暋崌懱傪宍惉偟偨屻丄NS4A偺N枛抂懁偺慳悈惈傾儈僲巁椞堟傪夘偟 偰嵶朎枌偲憡屳嶌梡偡傞偙偲偑帵嵈偝傟偰偄傞(12)丅
昅幰傜偺NS3僾儘僥傾乕僛扨撈偺峔憿偲丄Kim傜偑曬崘偟偨NS3僾儘僥傾乕僛 偲NS4A偺21偐傜39斣偺巆婎傪娷傓晹暘儁僾僠僪偲偺暋崌懱偺峔憿(5)傪斾妑 偡傞偲丄NS3僾儘僥傾乕僛扨撈偱偼丄偦偺N枛抂晹暘偼傎偳偗偰偄傞丅偦傟偵懳 偟暋崌懱拞偱偼丄偙偺NS4A偺儁僾僠僪嵔偲NS3僾儘僥傾乕僛偺A1偺儁僾僠僪嵔 側偳偲兝僔乕僩傪宍惉偟偰丄媴忬抈敀幙偺堦晹偲偟偰慻傒崬傑傟偰偄傞(5)丅NS3 僾儘僥傾乕僛扨撈偺応崌丄偦偺N枛抂晹埵偵偼丄Val-33丄Val-35丄Leu-44丄Leu-94丄 Val-107丄Leu-127丄Ala-111丄Val-113丄Pro-115丄Pro-142丄Leu-144丄Tyr-105(懁嵔)丄 Arg-109(庡嵔)偐傜側傞丄栺20x8侌偺峀偄慳悈椞堟偑宍惉偝傟傞丅嫽枴怺偄偙偲 偵丄NS4A晹暘偺側偄僾儘僥傾乕僛扨撈偺寢徎峔憿偱偼丄椬偁傞偄偼椉椬偵懚嵼偡 傞僾儘僥傾乕僛偺傎偳偗偨N枛晹暘偑丄偁偨偐傕NS4A儁僾僠僪偺擛偔丄偙偺慳悈 椞堟偲屳偄偵寢崌偟埨掕壔偟偰偄傞傛偆偵尒偊傞丅NS4A抈敀幙偼丄慳悈惈偺傾儈 僲巁偵晉傓攝楍傪桳偟偰偍傝丄NS3僾儘僥傾乕僛偺偙偺慳悈惈椞堟偲嫮偔憡屳嶌梡 偟丄屳偄偵埨掕壔偟偰妶惈傪憹戝偝偣偰偄傞偺偐傕偟傟側偄(4)丅
偍傢傝偵
C宆娞墛僂僀儖僗偺NS3偺僾儘僥傾乕僛椞堟偵偮偄偰丄怴偨偵夝柧偝傟偨寢徎峔 憿偵婎偯偄偰偦偺僾儘僥傾乕僛妶惈偲棫懱峔憿偲偺娭楢偵偮偄偰弎傋偰偒偨丅杮峺 慺偼丄婎幙偲偺憡屳嶌梡丄傑偨曗場巕偱偁傞NS4A偲偺憡屳嶌梡偐傜丄崱傑偱偵側 偄摿挜傪帩偮僙儕儞僾儘僥傾乕僛偱偁傞偙偲偑傢偐偭偨丅抈敀幙寢徎峔憿夝愅偱摼 傜傟偨NS3僾儘僥傾乕僛偺棫懱峔憿偼丄慾奞嵻偺憂弌偵戝偒側堦彆偵側傞傕偺偲峫 偊傜傟傞丅
偙偙偱摼傜傟偨峔憿偼NS3僾儘僥傾乕僛椞堟偵懳偡傞傕偺偱丄僂僀儖僗僎僲儉偵 僐乕僪偝傟偰偄傞揤慠宆峺慺偦偺傕偺偱偼側偄丅揤慠偱偼631傾儈僲巁巆婎偐傜側 傞NS3抈敀幙偲54傾儈僲巁巆婎偐傜側傞NS4A抈敀幙偑暋崌懱傪宍惉偟偰惗懱拞 偱婡擻偟偰偄傞偲峫偊傜傟傞丅NS3偺堦晹暘偱偁傞1-189傾儈僲巁偐傜側傞僾儘僥 傾乕僛椞堟偼丄偙偺揤慠宆偺NS3-4A暋崌懱偲偼丄婎幙偺擣幆丄憡屳嶌梡偵偍偄偰 婔暘側傝偲傕堎側傞壜擻惈偑斲掕偱偒側偄丅
嵟嬤丄NS3偺巆傝偺晹暘偱偁傞C枛抂懁2/3偺椞堟偵僐乕僪偝傟傞僿儕僇乕僛 椞堟偺寢徎峔憿偑曬崘偝傟偨(13)丅NS3抈敀幙偑帩偮慡偔堎側傞2偮偺峺慺妶惈 偑丄僂僀儖僗姶愼丒憹怋偺拞偱嬶懱揑偵偳偺傛偆偵婡擻偟偰偄傞偺偐偼嫽枴怺偄丅 偝傜偵丄HCV僂傿儖僗偺暋惢偵偍偄偰丄嵶朎撪偱嫟栶揑偵嶌梡偟偰偄傞偲峫偊傜傟 傞NS3-4A偺僿儕僇乕僛傗NTP傾乕僛妶惈偲丄NS5B偺RNA埶懚宆RNA億儕儊 儔乕僛妶惈偺憡屳嶌梡傗挷愡婡峔偺夝柧偑懸偨傟傞丅偙傟傜偺彅栤戣傪傂偲偮傂偲 偮夝寛偟偰偄偔側偐偐傜丄C宆娞墛偲偄偆擄昦傪崻帯偡傞怴偨側巺岥偑尒偊偰偔傞 傕偺偲怣偠偰偄傞丅
嵟屻偵丄偙偺尋媶惉壥偼丄Agouron Pharmaceuticals, Inc.偲偺嫟摨尋媶偵傛傝摼 傜傟偨傕偺偱偁傞丅傑偨丄恾偺嶌惢偲桳塿側彆尵傪偄偨偩偄偨姱栰夒巌丄敧楺岞 晇椉巵傪偼偠傔丄JT偺奆條偵姶幱偟傑偡丅嵟屻偵丄曻幩岝偺棙梡偵嫤椡偄偨偩偒 傑偟偨嶁晹抦暯巵傎偐PF偺僗僞僢僼偵姶幱偟傑偡丅
嶲峫暥專