
TARAプロジェクト報告書
平成9年10月29日
はじめに
我々の研究プロジェクトは機関誌「構造生物」を発行し、活動報告を詳しく記載している。従って、詳しい内容は資料1として添付した「構造生物」を参照し て頂きたい。
1. 本プロジェクトの目的と発足までの経過。
本プロジェクトの研究目的は産官学が一体となって、生物の個体維持と存続を機能的な側面から理解することである。即ち、最先端の技術開発を行いながら、 基礎として最も重要と考えられる関連物質の3次元構造をX線結晶解析法により決定し、得られた構造を基礎に生命体の制御機構を研究する。更に、コンピュー ターグラフィックスによる分子設計等を利用して、農業、医学、工業への利用研究の基盤作りも行う。
上記の目的を達成するためには、まず組織作りと経済的基盤を確立する必要がある。現在蛋白質のX線結晶解析には放射光利用が是非とも必要である。幸い筑 波大学は高エネルギー物理学研究所・放射光実験施設と極めて近距離にある。本プロジェクトは企業にとっても魅力のあるものと思われるので、産官学の共同研 究が成り立つ条件があると判断して本プロジェクトを組織した。また、本プロジェクトを研究内容に従って表1に示した9サブプロジェクト(以後班と略す)に 分け、各班に班長を置いて班の運営を効果的に進められるようにした。
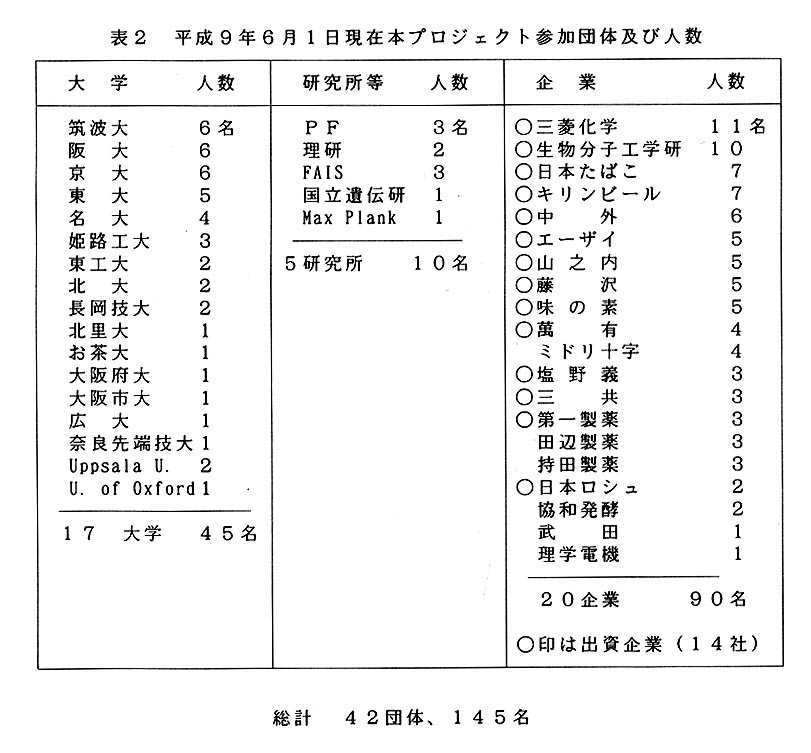
2. 本プロジェクトの組織と財政基盤。
本プロジェクトの構成を表2に示す。現在既に42団体145名から構成されている。
本プロジェクトの経費は企業からの寄附金とTARAからの補助金及びその他科研費など官からの補助金により賄っている。現在参加企業数は表2にあるように20社である。このうち14企業が寄付を行っている。
3. 第1班の活動
a)実験ステーションの建設
第1班の当面の任務は早急にTARA用実験ステーションを建設し平成8年5月から実験が開始できるよう整備することである。多くの方々の協力により計画 通りに建設が進み、平成8年5月15日よりテスト運転を開始し、5月22日よりTARA利用者の使用が開始された。
此のビームラインは輝度が大きく、カメラが使い易く、大型のイメージングプレート(IP)が使用できしかも精度も高いこと等により、極めて高い評価を得 ている。良い点を次に示す。
本プロジェクトスタート時点ではこの建設予定は無かったが、莫大なデータが TARA実験ステーションで得られ、それらを処理するのはTARA実験ステーション付近のみで行うことは極めて能率が落ちる。利用人口を本プロジェクトの メンバー数とすると145人であるが、実際には大学のメンバーは原則として研究室1人が本プロジェクトのメンバーに登録できる事にし、45名が登録されて いる。しかし実際の実験には院生や学生も来るので実験に来る人口は約4倍で大学からは180人程度である。そこでコンテナハウスの建設を行った。最初は 80m2の建設を行い、取りあえず電算機の一部を設置したがこれだけでは極端に不足であったため、現在は480m2にした。
c)CCD検出器を用いたX線テレビカメラの開発
計画通りCCD1個を用いるX線用検出器は平成8年度に完成した。しかし、これでは受光面積が不足なため、今後の発展性を考慮し、アレイ型の開発を行っ た。特に受光面の1/2をアルミ蒸着で遮光しバッファーとして使用することにより、露光中に読み出しが出来る検出器の開発を行うことにした。現在4個の CCDを2×2のアレイ型にし、3対1のテーパードオプティカルファイバーで受光面を拡大したものの開発を終了した。
4. 第2班の活動
a)各種電子計算機の導入
蛋白質結晶解析は数万~数百万の回折データを基に莫大な計算を行い、蛋白質の立体構造を求める。そのために電子計算機とそのソフトウエアが必要不可欠
である。既に購入し研究に供している電子計算機を次に示す:
b)各種プログラムの導入
既にインストールされたソフトウェアを次に示す:
WElS,CCP4,PROTElN,XTALVIEW,XPLOR,AUTOMR,DENZ0,0,TURB0-FRODO,MOLSCRIPT,Xsight,Quanta.
c)ネットワークの建設
TARA実験ステーションとTARAハウス間を光ファイバーで結び更にクラッカーなどに侵害されないようファイヤーウオールを取り付け外部と結ぶネットワ ークを完成させた。
d)TARA実験ステーションや電子計算機等の予約システム
本装置を利用するに当たりネットスケープを利用した予約システムを開発した。
5.第8班~第9班の活動
平成8年5月1日よりTARA実験ステーションの試運転が始まり、5月23日に本格的な利用が開始された。PFの入射器及びリングの大改装のため平成 8年12月16日より平成9年l l月まで放射光実験が停止した。従って実際に実験が出来たのは平成8年5月23日~7月21日及び10月26日~12月 15日の僅かな期間である。それにもかかわらず精力的な研究が行われ既にTARAに対する謝辞のある論文が37編報告(資料:論文リスト参照)されている。
a)第3班
①金属内包フラーレン電子Sc2@C84の電子密度1);坂田誠等はマキシマムエントロピー法を用い超精密解析を行っている。今回TARAとして行った研究を報告する。フラーレン化合物は、電子受容性など種々の側面から新物質としての関心が持たれている。特に、金属内包フラーレンは、超分子の典型とも言えるも ので、この種の分子の生成を自由にコントロールすることにより、全く新しい物質群の創製につながるものと期待されている。しかし、肝心の金属内包構造に関 しては、理論的予測や、間接的実験証拠による仮説の上に成り立っていた。最近、TARAにおける超精密解析の一環として進められてきたマキシマムエントロピ 一法により、金属原子をフラーレン・ケージ内に2個内包したSc2@C84の電子密度が、放射光X線回折実験データを使用し、求められた。その結果によると、2個のSc原子はC84ケージ中にケージ中心に対して、ほぼ対称に内包されていること、Scが2価の陽イオンとして存在することが分かった。また、フラー レン・ケージの対称性はD2dであること、さらには、C84ケージは内包された2個のSc原子との相互作用により、回転運動が殆ど凍結していることが分かった。 解析の結果得られた電子密度より計算された構造因子に基づく積分反射強度と観側値とは、R因子で2.1%と非常に良い一致を示した。
②2亜鉛インスリン結晶の超精密解析;坂部知平、坂部貴和子、佐々木教祐等は1978年に2亜鉛インスリン結晶の超精密解析を行い世界で初めて蛋白分子中 の水素原子を差フーリエ上で観察する事に成功した。その後 1.2Å 分解能のデ一夕を基に精密化を重ね1981年にはインスリン分子中、解離しない水素原子 の大部分の観察に成功した。例えばメチル基はNMRの測定では回転していると結論されるが、X線解析では3個の水素原子が回転方向へ延びた形ではっきり観 測できる、つまりメチル基はフリッピングモードの回転をしていることが判明した。その他多くの知見が得られた。更に1984年には2亜鉛インスリン結晶の 亜鉛のd-電子が差フーリエ上に観測される可能性が見付かった。即ち理論上推測される方向に電子密度が観測されたが距離的には理論値の2倍程の位置である 3)。これは測定データの分解能が不足しているためと説明出来るが証明にはならない。予備的な実験で2亜鉛インスリン結晶は分解能0.9Åまで反射が出るこ とが分かった。そこでインスリンの超精密解析用のデータを平成9年12月に測定する予定である。
b)第4班
呼吸は生体エネルギー産生の場として最も重要な生命現象の一つである。呼吸を行う電子伝達系は多数の構成成分より成るが、そのうち最も重要な機能を果た すタンパク質について、本プロジェクト班員により偉大な成果が得られた。
①ノーベル化学賞受賞者・H.Michel(マックスプランク研究所)のグループは、2.8Åの分解能で土壌細菌Paracoccus denitrificansのシトクロムc酸化酵 素の構造決定に成功し、呼吸のメカニズムを解明した。本酵素は4つのサブユニットから成るが、モノクローナル抗体FV断片と複合体を形成することにより結 晶化に成功した。4つのサブユニットのうち最も重要な活性中心をもつのはサブユニットⅠで、12の膜貫通セグメントを含み、また2つのヘムa(ヘムa、ヘ ムa3)と1個の銅原子(CuB)をもつ。ヘムa 3とCuBは2核中心を形成し、ここで分子状酸素は水へと還元される。シトクロムcからの電子はサブユニ ットⅡにある銅(CuA)に先ず渡され、ヘムaを中継して2核中心に到達する。 本酵素には2つのプロトン移動経路が存在する。すなわち一つは分子状酸素と反応して水を作り、もう一方はプロトンを膜の外に汲み出すプロトンポンプとして 働く経路である。プロトンポンプの作用で膜の内外にプロトンによる電気化学的ポテンシャルが形成され、それがATP生成の駆動力となるが、構造解析の結果 をもとにプロトンポンプのメカニズムを解明した。
②同じくH.Michelのグループは、呼吸鎖電子伝達系のシトクロムb c1 複合体中の一成分である、特異な鉄・イオウクラスター,'Rieske’の構造決定にも成功 した。シトクロムb c1 複合体をトリプシン消化し、完全な還元型Rieskeタンパク断片を得、MADにより1.5Åの分解能で構造決定を行った。2個の鉄に 配位するリガンドはシステインとヒスチジンが2個づつであり、4個全てがシステインである従来のフェレドキシンとは異なっていた。得られた構造をもとにシ トクロムb c1複合体によるキノール酸化のメカニズムを示した。
③化学合成独立栄養生物である硝化細菌Nitrosomonas europaeaは、アンモニア を亜硝酸へ酸化することでエネルギーを得ている。 アンモニアはアンモニアモノオキシゲナーゼとヒドロキシルアミン酸化酵素(HAO)により亜硝酸に酸化されるが、エネルギー(ATP)生成に関わるのはHAOのみである。このエネル ギー産生に関わるHAOの構造を2.8Åの分解能で決定した。HAOはヘムcを8個もつサブユニットの3量体で、合計24個もヘムを有する。各ヘムは分子 中央の底に集まり、サブユニットあたり4つのクラスターを形成している。3量体中のヘムクラスターは整列して環をなし、入口と出口を形成している。8個の ヘムのうちの一つは一酸化炭素(CO)結合能を有し、その吸収スペクトルからP460と呼ばれる。P460は入口に位置し、基質ヒドロキシルアミンから2 電子を同時に受け取る。ヘムに2電子が直接渡される現象は、全く新しい生体電子伝達機構である。電子の出口は2箇所あり、一方の出口はエネルギー産生のた めの電子伝達系、シトクロムc 554、c 552および末端酸化酵素へと連結していると思われる。この種の(化学合成独立栄養)細菌は成育が悪く、結晶構造解析が できる程のタンパク量を確保することが困難であった。ヘムを24個ももつ本酵素の構造は、Nature Structural Biology掲載誌の表紙を飾った。
c)第5班
①C型肝炎ウイルスの複製において、ポリタンパクプロセシングの最後のステップは非構造タンパク質3(NS3)のアミノ末端側3分の1を構成するウイルス
自身のタンパク質分解酵素によりなされる。このNS3タンパク質の構造を2.4Å分解能で決定した。NS3は二つのβバレルと触媒トリアード、His-57,
Asp-81,Ser-139、より成るトリプシン様構造をとっていた。また基質結合部位の構造は切断特異性によく一致していた。一方、亜鉛結合部位の構造や、分子表
面疎水パッチヘの結合により隣接分子と相互作用する長いアミノ末端構造の存在など、新しい発見もされた。
この結果は、NatureやScience誌を凌ぐインパクトファクターを誇るCellに掲載された。
d)第6班
非常に多くの酵素・タンパク質の構造解析、反応機構などが明かにされた。
①マウス肺カルボニル還元酵素(MLCR)は短鎖脱水素酵素/還元酵素(SDR)ファミリーに属するが、基質2-プロパノールおよびNADPHとの3者複合
体の構造決定を行った。本酵素がNADHとNADPHをどのように見分けているのか不明であったが、構造解析によりそれが明かとなった。即ちSDRファミ
リーの酵素では、NADHおよびNADPH分子の唯一の違いであるリボースの2'-部位を識別し、NADHに特異的な場合は負荷電アミノ酸残基が、NADP
Hに特異的な場合は正荷電アミノ酸残基がそれぞれ結合部位となっていると結論された。
ダイオキシンやPCB(ポリ塩化ビフェニル)などの塩化フェニル化合物は、毒性の高い環境汚染物質として問題となっている。PCBを分解できるPseudomonas
属細菌の分解系成分であるBphCの構造決定を行った。この酵素は非ヘム鉄を合み、芳香環エキストラジオール型開裂を行うジオキシゲナーゼであるが、
このタイプのオキシゲナーゼとして最初の構造決定である。
大腸菌の7α-ヒドロキシステロイド脱水素酵素の2者複合体(酵素・NAD+)および3者複合体(酵素・NADH・7-オキソグリコケノコール酸)の構造決
定を行った。本酵素もSDRファミリーに属するが、これら複合体の構造決定により本ファミリー酵素の普遍的な触媒メカニズムを明かにした。
②大腸菌のγ-グルタミルトランスペプチダーゼの構造解析を3.0Å分解能で行い、プレリミナリーな結果を得た。
③カビの脱窒に関与するシトクロムP450norは、P450スーパーファミリ一に属しながら一酸化窒素(NO)の還元という特異な反応を行い、さらにNADH から直接電子を受け取るなど、興味深い反応機構をもつことで注目される。 本P450の休止酸化型およびCO型の構造決定を行った。特異な反応機構にも関わらずP450norの構造は他のP450と非常によく似ていた。一方、ヘム 遠位ポケットに大きなオープンスペースが存在し、これはNADHのアクセスチャネルであると予想した。2種の型の比較から、プロトン供給系としての水分子 ・アミノ酸側鎖水素結合ネットワークの存在が示唆された。 さらに細密な構造解析を目的に、良好な単結晶の作成に成功した。 同じカビから、フラビン蛋白質とヘム蛋白質の融合タンパクであるフラボヘモグロビンを単離した。生理機能は不明であるが構造に興味がもたれ、結晶を作成 する予定である。
④中度通性好熱菌Bacillus coagulansの3-イソプロピルリンゴ酸脱水素酵素の結晶を3種得た。そのうちの一つが結晶解析に適していた。
リポアミド脱水素酵素はα-ケト酸脱水素酵素複合体に普遍的に見出される構成成分である。この酵素を酵母より高度に生成し、プレリミナリーな構造解析の
結果を得た。
⑤放線菌Streptomyces olivaceoviridisのβ-キシラナーゼを構造決定した。 チューリップ由来の除草剤分解酵素アリルアシルアミダーゼについて、結晶の作成および2.5Å分解能X線データ測定の結果を報告した。
⑥好酸好熱古細菌Sulfolobus由来のフェレドキシンの構造決定を2.0Å分解能で行った。古細菌フェレドキシンに特徴的な構造として、4つのアミノ酸残基
が亜鉛原子に配位した四面体構造の存在を初めて明かにした。
細菌Bacillus cereusオリゴ-1,6-グルコシダーゼの構造解析を2.0Å分解能で行った。構造は3つのドメインより成り、触媒部位であるAsp-199,Glu-
255,Asp-329の位置も特定された。さらに同種酵素の一次配列の比較から予想されていた、熱安定性に重要な21のプロリン置換部位が明かとなった。また、ル
ープ中よりもβ-夕ーンおよびα-ヘリックス中のプロリン置換部位の方が熱安定性に重要であることが明かとなった。
⑦細菌Paracoccus denitrificans由来シャペロニン-60(cpn60)を結晶 化し、3.2Å分解能の回折データを得た。
⑧Arthromyces ramosusペルオキシダーゼとベンズヒドロキサミン酸(BHA)との複合体の構造解析を1.6Å分解能で行い、BHAの結合様式を細部にわた り明かにした。
⑨放線菌Streptomyces caespitosusの亜鉛エンドペプチダーゼは、132アミノ酸より成る、これまで知られている最も小さいプロテアーゼである。本酵素は、 亜鉛結合配列HEXXHをもつものの、他の亜鉛メタロエンドプロテアーゼとの一次配列相同性は低い。しかしながら全体の構造は、亜鉛メタロエンドプロテア ーゼスーパーファミリーとよく似ていた。亜鉛原子はHEXXH配列中の2個のヒスチジン、アスパラギン酸、および水分子と配位し、4面体構造を形成してい た。従って本酵素は、亜鉛結合配列HEXXHXXGXXDをもつ新サブファミリーに属する。
e)第7班
酵素の反応機構を研究する方法は種々あるが反応の過程を8次元構造の変化として見る方法は結晶解析のみである。水溶液中に於ける多くの酵素の反応速度は ミリ秒~秒の領域にある。この速度で構造変化を追究するためには放射光を用い たラウエ法しかない。TARA用ビームラインBL6Bは時分割ラウエ法には使 用できない。従ってこの研究は厳密に言うとTARAとしては研究できないこと になるが、しかし坂部知平が代表として行われている重点領域の研究には本プロ ジェクトの班員が多数参加しているので、その結果を極簡単に報告する。
①水平方向にIPカセットを移動させながら幾度も露光(5ミリ秒以上)を繰り 返すことで横軸を時間軸とする時分割ラウエカメラを製作し平成7年度にBL18Bに設置した。
②時間分割ラウエ法適用のための結晶化法の開発、特殊条件化に於ける回折強度 測定法の研究、ラウエデータによる迅速精密解析法の研究等を行い多大な成果 をあげた。
③時分割ラウエ実験で最も大切なことは如何にして反応を一斉にスタートさせる かということである。トリガーとしてはレーザーやフラッシュランプ、T-ジ ャンプ、p H-ジャンプ等が考えられ、全てについてサーベイが行われた。予 め蛋白結晶中に基質のケージ化合物を入れておき光照射で反応を開始する方法 が有力と考えられたが、多くの困難があり今後更に多くの研究を行う必要があ る。それでもグルタチオン合成酵素はこの方法を用いて反応の中間状態を時間 分割して観察できた。
④最も大きな発見は溶液中でミリ秒オーダーの反応速度を持つ酵素が結晶中では 秒から分オーダーの反応速度になることである。要するに結晶中では秒~分オ ーダーの測定をすれば溶液中のミリ秒オーダーの反応が追えるという事を見い 出した。この事により小さな結晶を用いれば、フローセルを用い基質を結晶に 染み込ませながら反応を測定する可能性も出てきた。この場合ラウエ法に頼ら なくても良く、TARA実験ステーションでの研究も可能である。
f)第8班
①多くのアルキル化塩基をDNAより除く大腸菌3-メチルアデニングリコシラー
ゼⅠの構造を2.3Å分解能で決定した。本酵素は、真核生物のTATAボック
ス結合タンパク質の半分と類似性をもつα+βフォールドドメイン1つと、大腸
菌エンドヌクレアーゼⅢによく似たα-ヘリックスより成るドメイン2つより成
る。活性中心は2つのヘリカルドメインの間のクレフトに存在し、また二重鎖D
NAから標的塩基を酵素が放り込むことが変異タンパク質およびモデル構築の検
討により示唆された。活性中心の構造は、広い基質特異性と単純なNグリコシラ
ーゼ活性を示唆した。
本論文もCellに掲載された。
②発生分化を研究する上での良いモデル生物である原生生物の細胞性粘菌を用い て、DNAトポイソメラーゼⅡ(Top II)が生物の発生・分化にどのように関 与しているかを調べた。本酵素の遺伝子をクローン化し、作成した抗体を用いた 実験からTop IIがミトコンドリアに局在し、またそのN末端にミトコンドリア への移行シグナルが存在することをTop IIタンパク質として初めて明かにした。 また同じ細胞性粘菌のミトコンドリアDNAのグループ1イントロン(リボザイ ムの一種)中に存在するorfの構造と機能を調べた。調べたorfの一つは、 大腸菌で発現させると一本鎖のみの切断を行うDNAエンドヌクレアーゼ活性を もっていた。
g)第9班
①T4リゾチーム活性中心のスレオニン26をヒスチジンに変換すると、反応産 物がα-からβ-アノマーヘと変わった。野生型と変異酵素の比較から、それぞ れの反応機構を推定した。また野生型T4リゾチームと卵白リゾチームとは共通 の祖先から派生したと思われるのに、異なるメカニズムをもつことが明かとなっ た。
②蛇毒由来の血液凝固IX因子およびX因子結合タンパク質について、2.5Å 分解能で構造解析を行った。解析の結果、サブユニット中央のループがドメイン スワッピングしたユニークな二量体構造をしていることが明かとなった。
③精巣上体における精子の運動能、受精能の獲得の分子機構を解明する目的で、 数種の精巣上体分泌タンパク質の働きを明かにしてきた。それらのうち、以下の 2種類のタンパク質結晶構造解析を試みている。ブタ135kDa-α-D-マンノシダー ゼは、受精の際、卵と接触する精子頭部の赤道帯と先端部に結合することが明か となった。精子の受精能の獲得を担う分子であると考えられる。一方マウスでは 精子形成との関連性も示唆された。15kDa-コレステロール結合タンパク質は、精 巣上体の頭部後半から体部にかけて分泌され、コレステロールを強く結合し、精 子細胞膜から抜き取る作用をもつことが明かとなった。成熟に伴う精子細胞膜の 質的変化を担うと思われる。これらのタンパク質について、既にcDNA遺伝子 共にクローニングし、塩基配列も決定している。
④造血細胞プロスタグランディンD合成酵素は、免疫系やマスト細胞における
プロスタノイドDおよびJ系列合成のキーエンザイムである。ラット酵素の
cDNAを単離し、その組み換え酵素を結晶化、グルタチオンとの複合体の構造
を2.3Å分解能で決定した。本酵素は脊椎動物由来のシグマクラスのグルタチ
オン-S-トランスフェラーゼ(GTS)の最初のメンバーであり、他のGTSフ
ァミリーメンバーには見られない顕著なクレフト構造の活性中心を形成していた。
そのクレフトの特異な3次元構造は、PGH2からPGD2への特異的異性化に
対応する基質結合様式と触媒機構をうまく説明可能であった。
この結果はCellに掲載された。