1丏偼偠傔偵
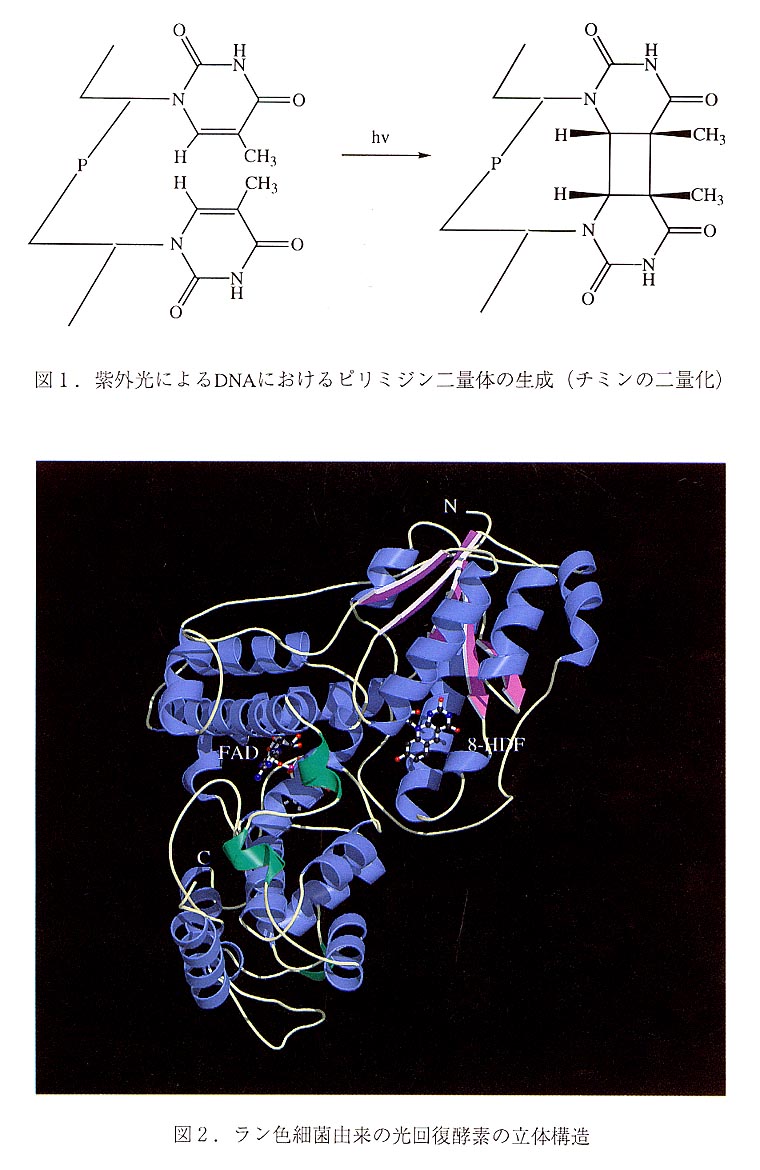
丂DNA偼丆擬丆巼奜慄丆僗僩儗僗摍偱梕堈偵曄堎傪庴偗傞丏偙偺條側曄堎(壔 妛曄壔)傪偆偗偨DNA偼惗懺宯偺堚揱忣曬堐帩偵巟忈傪棃偡嫲傟偑偁傞偑丆偙偆偟 偨懝彎DNA傪廋暅偡傞峺慺孮偑惗懱撪偵偼懚嵼偡傞丏偙傟傜偼DNA廋暅峺慺偲屇偽 傟偰偄傞丏岝夞暅峺慺(photolyase)偼DNA廋暅峺慺偺堦偮偱丆巼奜慄偵傛偭偰DNA 偵惗偠傞懝彎偺堦偮偱偁傞僺儕儈僕儞擇検懱(僠儈儞偺擇検壔丆恾1)偵寢崌偟丆 偦偺屻偵媧廂偡傞岝偺僄僱儖僊乕傪梡偄偰尦偺扨検懱偵廋暅偡傞丏偙偺DNA廋暅偼 岝夞暅偲屇偽傟傞丏
丂儔儞怓嵶嬠桼棃偺岝夞暅峺慺偼丆暘巕検栺5枩偺傾億僞儞僷僋幙偲僼儔價儞 崪奿傪傕偮2庬偺曗寚暘巕偐傜側偭偰偄傞(1). 岝夞暅峺慺偼DNA偵惗偠傞懝彎DNA 偺僺儕儈僕儞擇検懱偵寢崌偟偨屻丆媧廂偟偨岝偺僄僱儖僊乕傪僩儕僈偲偟偰擇検懱 偐傜扨検懱傊偺廋暅傪峴偆丏嫽枴怺偄偙偲偵廋暅帪偵梡偄傞攇挿偑丆挿攇挿宆偺岝 夞暅峺慺(乣440nm傪媧廂)偲抁攇挿宆偺傕偺(乣380nm)偵暘椶偡傞偙偲偑偱偒傞丏 杮尋媶偱懳徾偲偟偨儔儞怓嵶嬠桼棃偺傕偺偼挿攇挿宆(2)丆堦曽丆戝挵嬠桼棃偺傕偺 偼抁攇挿宆偵懏偡傞(3). 偙傟偼丆2庬偺曗寚暘巕僼儔價儞偺峔憿偺堘偄偵婲場偟偰 偄傞丏偳偪傜偺峺慺傕娨尦偝傟偨FAD傪戞1偺曗寚暘巕偲偟偰嫟捠偵傕偭偰偄傞偑丆 戞2偺曗寚暘巕偺庬椶偑堎側傞偙偲偵傛傝丆挿丆抁攇挿宆偺堘偄偑惗偠偰偄傞丏偡 側傢偪丆挿攇挿宆偱偼僨傾僓僼儔價儞宆曗寚暘巕傪丆抁攇挿宆偱偼僾僥儕儞宆曗寚 暘巕傪桳偟偰偄傞丏偙偺戞2偺曗寚暘巕偼岝傪媧廂偟偰丆偦偺僄僱儖僊乕傪娨尦宆 FAD偵搉偡廤岝曗寚暘巕偱偁傝丆偙偺僄僱儖僊乕傪庴偗偰怗攠曗寚暘巕偱偁傞FAD 偵傛傞僺儕儈僕儞擇検懱偺廋暅偑峴傢傟傞丏
丂嵟嬤丆僾僥儕儞宆偺廤岝曗寚暘巕傪帩偮戝挵嬠桼棃偺峺慺偺寢徎峔憿偑 Deisenhofer偺僌儖乕僾偵傛偭偰夝柧偝傟偨偑(4)丆堦曽丆変乆偼僨傾僓僼儔價儞宆曗 寚暘巕傪帩偮儔儞怓嵶嬠桼棃偺峺慺偺寢徎峔憿夝愅偵惉岟偟偨(5). 偦偺寢壥丆嬃偔 傋偒偙偲偵丆慡懱偺僼僅乕儖僨傿儞僌偵偼崅偄嫟捠惈偑偁傞偵傕偐偐傢傜偢丆廤岝 曗寚暘巕偺寢崌晹埵偑慡偔堎側偭偰偄傞偙偲傪尒偄弌偟偨丏偙傟偼廤岝曗寚暘巕偐 傜怗攠曗寚暘巕(FAD)僿偺僄僱儖僊乕堏摦偑椉峺慺偱堎側偭偰偄傞揰偐傜嬌傔偰嫽 枴怺偄丏
2丏儔儞怓嵶嬠桼棃岝夞暅峺慺偺X慄峔憿夝愅
丂儔儞怓嵶嬠桼棃偺岝夞暅峺慺偺寢徎偼丆棸巁傾儞儌僯僂儉傪捑揳嵻偲偟偰忲 婥奼嶶朄偵傛傝摼傜傟偨丏拞惈晅嬤偺pH偱摼傜傟偨寢徎偼丆拰忬寢徎偑廤傑偭偨僋 儔僗僞乕忬懺偱偁偭偨偑(6)丆EDTA傪揧壛偡傞偙偲偵傛傝丆榋妏拰忬偺扨撈偺寢徎偲 側偭偨(7). 偄偢傟偺寢徎傕PF偺BL6A僺乕儉儔僀儞偵偍偄偰嫄戝暘巕梡儚僀僛儞儀 儖僋僇儊儔(嶁晹僇儊儔)傪梡偄偰X慄夞愜幚尡傪峴偆偙偲偑偱偒偨偑丆僋儔僗僞 堦忬拰忬寢徎(僋儔僗僞乕偐傜愗傝弌偟偰梡偄偨)偑1.8侌暘夝擻傪梌偊偨偺偵懳偟 偰丆榋妏拰忬偺寢徎偼3.0侌暘夝擻掱搙偺夞愜揰偟偐娤應偡傞偙偲偑偱偒側偐偭偨丏 偟偨偑偭偰丆拰忬寢徎傪梡偄偰峔憿夝愅傪峴偭偨丏偙偺拰忬寢徎偼丆惓曽徎宯偵懏 偟丆嬻娫孮P43212丆a=b=91.17侌丆c=135.54侌偱丆旕懳徧扨埵拞偵堦暘巕偺岝夞暅峺 慺傪娷傫偱偄偨丏夞愜嫮搙僨乕僞偺應掕傕PF偱嶁晹僇儊儔傪梡偄偰峴偭偨丏3庬偺 廳尨巕傪梡偄偨摱尨巕摨宆抲姺朄偵傛傝丆1.8侌暘夝擻偺寢徎峔憿夝愅傪峴偆偙偲偑 偱偒偨丏
3丏儔儞怓嵶嬠桼棃岝夞暅峺慺偺棫懱峔憿
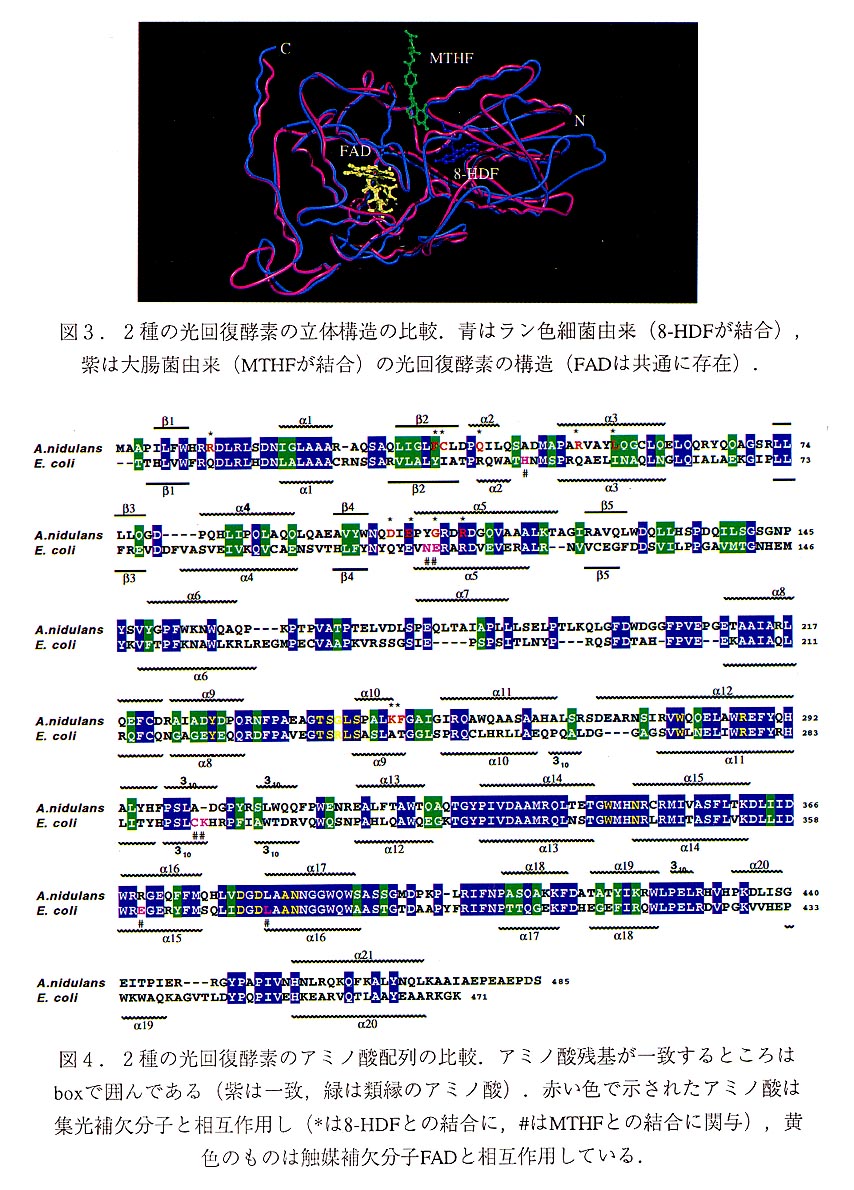
丂儔儞怓嵶嬠桼棃偺岝夞暅峺慺偺慡懱峔憿傪恾2偵帵偡丏暘巕偼擇偮偺僪儊僀 儞偵暘偐傟傞丏N枛抂懁偼偄傢備傞Rossmannfold傪偲偭偰偄傞兛/兝僪儊僀儞偱偁傝丆 堦曽丆C枛抂懁偵懡偔偺僿儕僢僋僗偐傜側傞僿儕僇儖僪儊僀儞偑埵抲偟偰偄傞丏兛11 僪儊僀儞偵偼丆廤岝曗寚暘巕偱偁傞8-僸僪儘僉僔-5-僨傾僓僼儔價儞(8-HDF)偑丆 堦曽丆僿儕僇儖僪儊僀儞偵偼怗攠曗寚暘巕偱偁傞FAD偑寢崌偟偰偄偨丏暘巕慡懱偺 僼僅乕儖僨傿儞僌偼愭偵寛掕偝傟偨戝挵嬠桼棃偺岝夞暅峺慺偲旕忢偵椙偔帡偰偍傝 (恾3)丆椉幰偺C兛扽慺偺r.m.s deviation偼2.2侌偱偁偭偨丏棫懱峔憿忋偺堘偄偼儔儞怓嵶嬠桼棃偺岝夞暅峺慺偵偺傒丆N抂懁偺兛/兝峔憿偵晅悘偟偰傕偆堦杮兛-僿儕僢僋僗偑懚嵼偟偰偄傞偙偲偱偁偭偨丏
丂恾4偵偼儔儞怓嵶嬠桼棃丆戝挵嬠桼棃偺岝夞暅峺慺偺傾儈僲巁攝楍傪帵偟偰 偁傞丏棫懱峔憿忋偺憡摨惈偲憡傑偭偰2偮偺峺慺偺堦師峔憿偼椙偔帡偰偍傝丆39% 偑摨堦巆婎丆13%偑惈幙椶帡偺巆婎偱偁偭偨丏
丂妶惈敪尰偵昁梫側曗寚暘巕偱偁傞FAD偼丆偄偢傟偺岝夞暅峺慺偱傕傎偲傫偳 摨堦偺埵抲偵懚嵼偟(恾3)丆2偮偺岝夞暅峺慺偺FAD暘巕偱偺r.m.s. deviation偼 0.24侌偱偁偭偨丏傑偨丆FAD暘巕偲憡屳嶌梡偟偰偙傟傪曐帩偡傞傾儈僲巁巆婎傕椉幰 偺峺慺偱旕忢偵傛偔堦抳偟偰偄偨丏偟偐偟側偑傜丆兛/兝僪儊僀儞偺廤岝曗寚暘巕偼丆 埲壓偵帵偡傛偆偵丆椉幰偱慡偔堎側偭偨埵抲偵寢崌偟偰偄偨丏
4丏廤岝曗寚暘巕偺擣幆
丂岝夞暅峺慺偺廤岝曗寚暘巕偼媧廂偟偨岝僄僱儖僊乕傪怗攠曗寚暘巕偱偁傞 FAD偵揱払偡傞丏怗攠曗寚暘巕偵傛傞僺儕儈僕儞擇検懱偺廋暅斀墳偵偼偙偺僄僱儖 僊乕偑昁梫偱偁傞丏愭偵帵偟偨傛偆偵丆儔儞怓嵶嬠桼棃偺岝夞暅峺慺偼廤岝曗寚暘 巕偲偟偰8-HDF傪丆戝挵嬠桼棃偺岝夞暅峺慺偼僾僥儕儞宆曗寚暘巕偱偁傞5,10-儊僥 僯儖僥僩儔僴僀僪儘梩巁(MTHF)傪桳偟偰偄傞丏儔儞怓嵶嬠桼棃偺岝夞暅峺慺偺寢徎 峔憿偑柧傜偐偲側偭偨偙偲偱丆2庬偺岝夞暅峺慺偵偍偗傞廤岝曗寚暘巕丆8-HDF(儔 儞怓嵶嬠)側傜傃偵MrHF(戝挵嬠)偺寢崌埵抲偼丆慡偔堎側偭偰偄傞偙偲偑傢偐 偭偨丏偄偢傟傕N枛偺兛11僪儊僀儞偵寢崌偟偰偄傞偑丆8-HDF偼MrHF偵斾傋偰12侌傕 暘巕偺撪懁偵擖傝崬傫偩埵抲偵偁傞丏偙偺兛/兝僪儊僀儞偺僼僅乕儖僨傿儞僌偼2偮偺 峺慺偱傎偲傫偳堦抳偟偰偍傝(恾5)丆偙偺晹暘偺傾儈僲巁攝楍偵偮偄偰傕兝1偐傜 兛6傑偱旕忢偵憡摨惈偑崅偄(恾4)丏偡側傢偪丆嶰師尦峔憿偑傎偲傫偳摨偠偱丆偟 偐傕傾儈僲巁巆婎偺憡摨惈偑嬌傔偰崅偄偵傕偐偐傢傜偢丆椉幰偦傟偧傟偱廤岝曗寚 暘巕偲憡屳嶌梡偡傞傾儈僲巁傪彮偟偢偮曄偊傞偙偲偵傛傝丆堎側傞廤岝曗寚暘巕傪 擣幆偟偰偄傞偙偲偑帵偝傟偨偺偱偁傞丏忋偵弎傋偨廤岝曗寚暘巕偲憡屳嶌梡偡傞傾 儈僲巁巆婎傪嬶懱揑偵恾6偱帵偡偲丆偨偲偊偽戝挵嬠偺MTHF偺寢崌偵娭梌偡傞 His44偼丆儔儞怓嵶嬠偱偼Ala45偲側偭偰偍傝丆8-HDF偑寢崌偡傞偨傔偺嬻娫傪梌偊偰 偄傞丏Phe80傕戝挵嬠偱偼8-HDF寢崌僒僀僩傪嵡偄偱偄傞偑丆儔儞怓嵶嬠偱偼偙偺晅 嬤偺傾儈僲巁偑寚懝偟偰偄傞丏Gly106偼8-HDF寢崌僒僀僩偺僗儁乕僗傪梌偊傞偺偵 偪傚偆偳椙偄偑丆戝挵嬠偱偼MrHF偲偺寢崌偵昁恵偺傾儈僲巁偱偁傞Glu109偵抲偒 姺偊傜傟偰偄傞丏儔儞怓嵶嬠偺Lys248傕8-HDF偲悈慺寢崌傪偡傞廳梫側巆婎偱偁傞 偑丆戝挵嬠偱偼Ala242偲側偭偰偄傞丏媡偵戝挵嬠偺Lys293偼MTHF偲偺悈慺寢崌偵昁 恵偱偁傞偑儔儞怓嵶嬠偱偼寚懝偟偰偄傞丏
丂戝挵嬠偼偦偺惗妶娐撪偱偼8-HDF傪惗嶻偡傞偙偲偼偱偒側偄丏堦曽丆儔儞怓 嵶嬠偼8-HDF傪帩偮偙偲偵傛偭偰丆傛傝挿搉挿懁偺岝傪廤岝偡傞偙偲偑偱偒偨丏暘 巕恑壔揑偵峫偊傞偲丆儔儞怓嵶嬠偑傛傝挿攇挿偺岝傪妉摼偡傞偨傔偵杮棃MrHF偑 寢崌偟偰偄偨晹暘偺傾儈僲巁傪彮偟偢偮曄堎偝偣偰恑壔偟丆8-HDF偺寢崌僒僀僩傪 帩偮偙偲偑偱偒偨傕偺偲巚傢傟傞丏
丂儔儞怓嵶嬠桼棃偺岝夞暅峺慺偺寢徎峔憿傪戝挵嬠桼棃偺峺慺偲偺峔憿傪斾妑 偡傞偙偲偵傛偭偰丆椉僞儞僷僋幙偺廤岝曗寚暘巕偺擣幆婡峔傪棟夝偡傞偙偲偑偱偒 偨丏旕忢偵傛偔曐懚偝傟偨堦師峔憿偲摨堦偺僼僅乕儖僨傿儞僌傪帩偭偨2偮偺僞儞 僷僋幙偵偍偄偰丆偦偺寢崌晹暘偺傾儈僲巁巆婎傪曄堎偝偣偰庬椶偺堎側傞曗寚暘巕 傪曐帩偟偰偄傞椺偼丆崱傑偱偵妋擣偝傟偨偙偲偑側偔丆峔憿惗暔妛忋旕忢偵嫽枴怺 偄偲巚傢傟傞丏
5丏偍傢傝偵
丂杮尋媶偼搶杒戝妛壛楊堛妛尋媶強埨堜柧嫵庼丆僆儔儞僟丆儘僢僥儖僟儉丒 僄儔僗儉僗戝妛偺A丏P丏M丏 Eker攷巑偲偺嫟摨尋媶偱丆嫗搒戝妛戝妛堾棟妛尋媶壢壔 妛愱峌偺惗暔峔憿壔妛尋媶幒(搶嫗岺嬈戝妛帒尮壔妛尋媶強惗暔帒尮晹栧偐傜偺宲 懕)偵偍偄偰峴傢傟偨傕偺偱偁傞丏
丂偙偺尋媶傪峴偆偵偁偨傝X慄夞愜幚尡偱戝偒側偛彆椡傪偄偨偩偄偨崅僄僱儖 僊乕壛懍婍尋媶婡峔丆暔幙峔憿壢妛尋媶強(曻幩岝尋媶巤愝)丆嶁晹抦暯柤梍嫵庼丆 拞愳撝巎攷巑(尰杒戝堾棟)丆搉绮怣媣攷巑丆楅栘庣攷巑偵屼楃怽偟忋偘偨偄丏
嶲峫暥專