偼偠傔偵
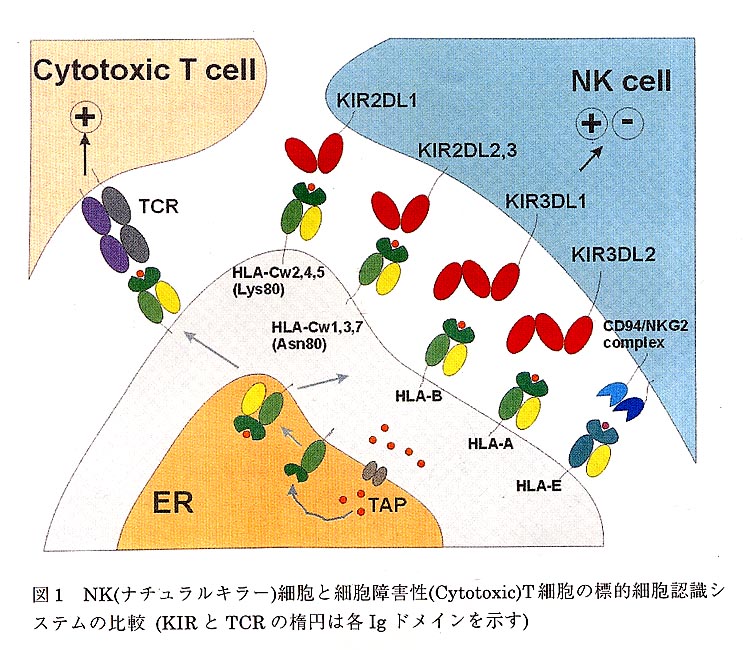
柶塽宯偵偼弶婜偺姶愼杊屼婡峔偱偁傞帺慠柶塽偲峈尨摿堎揑側妉摼柶塽偺2庬椶偺僔僗僥儉偑偁傞丅妉摼柶塽傪扴偆儕儞僷媴偺1偮偱偁傞嵶朎忈奞惈T嵶朎偱偼丄桳妀嵶朎昞柺偺庡梫慻怐揔崌惈峈尨暋崌懱(MHC)僋儔僗I(僸僩偱偼僸僩敀寣媴峈尨丄HLA-A丄HlMA-B丄HLA-C偲屇偽傟傞)偵寢崌偟偰偄傞僂僀儖僗傗庮釃桼棃偺旕帺屓儁僾僠僪傪峈尨摿堎揑T嵶朎儗僙僾僞乕(TCR)偑擣幆偟丄惓偺僔僌僫儖傪憲傝丄MHC峉懇惈嵶朎忈奞妶惈傪帵偡(恾1)丅懠曽丄帺慠柶塽偵娭傢傞摨偠偔儕儞僷媴偱戝宆偺梓棻傪傕偮僫僠儏儔儖僉儔乕(natural killer丄NK)嵶朎偼僂僀儖僗姶愼嵶朎傗庮釃嵶朎側偳傪峈尨摿堎揑巋寖側偟偵嶦偟偨傝丄條乆側僒僀僩僇僀儞傪暘斿偡傞丅廬棃丄NK嵶朎偺嵶朎忈奞妶惈偼MHC旕峉懇惈偱偁傞偲峫偊傜傟偰偒偨偑丄嵟嬤偄偔偮偐偺MHC僋儔僗I暘巕偵摿堎揑偵寢崌偡傞NK嵶朎儗僙僾僞乕暘巕孮偑敪尒偝傟丄T嵶朎偲摨偠偔MHC峉懇惈傪帵偡偙偲偑傢偐偭偰偒偨丅偙傟傜偺NK嵶朎儗僙僾僞乕偼丄T嵶朎儗僙僾僞乕偲偼懳徠揑偵MHC僋儔僗I傪擣幆偟偰晧偺僔僌僫儖傪憲傝丄寢壥偲偟偰惓忢偵MHC僋儔僗I暘巕傪敪尰偟偰偄傞嵶朎偼嶦偝側偄丅媡偵丄MHC僋儔僗I暘巕傪敪尰偟偰偄側偄堎忢側嵶朎偼T嵶朎偵偼擣幆偝傟側偄偑丄NK嵶朎偵傛傝嶦偝傟傞丅T嵶朎偲NK嵶朎偼偙偺揰偵偍偄偰憡曗揑栶妱傪扴偭偰偄傞偲巚傢傟傞(恾1)丅僸僩偱偼MHC僋儔僗I摿堎揑側NK嵶朎儗僙僾僞乕孮偑2庬椶偁傝丄1偮偼C-type儗僋僠儞僼傽儈儕乕偵懏偡傞CD941NKG2僿僥儘暋崌懱僼傽儈儕乕偱偁傝丄懠曽偼柶塽僌儘僽儕儞(Ig)僼傽儈儕乕偵懏偡傞僸僩Kiler cell immunog1obulin (Ig)-like receptor(KIR丄Killer cell inhibitory receptor)僼傽儈儕乕偲僸僩Ig-like transcript(ILT)/leukocyto Ig-like receptor(LIR)僼傽儈儕乕偱偁傞丅CD94/NKG2暋崌懱偼non-classical MHC偱偁傞HLA-E傪擣幆偡傞偺偵懳偟偰丄KIR傗ILT偼MHC僋儔僗I傪擣幆偡傞(堦晹偺ILT偼non-classical MHC偱偁傞HLA-G傕擣幆偡傞傛偆偱偁傞)丅偄偢傟偺僼傽儈儕乕偲傕嫽枴怺偄偙偲偵immunoreceptor tyrosine丂based inhibitory motif(ITIM乺偺懚嵼偡傞挿偄嵶朎撪僪儊僀儞傪傕偮傕偺偲ITIM傪帩偨側偄抁偄嵶朎撪僪儊僀儞偺傕偺偑偁傞丅抁偄曽偼immunoreceptor tyrosine-based activating motif(ITAM)傪傕偮DAP12側偳偵寢崌偡傞丅偦傟備偊丄偙偺儗僙僾僞乕孮偼NK嵶朎偺妶惈壔偲偦偺慾奞偺椉曽偵娭梌偟丄偦偺僶儔儞僗偱NK嵶朎偺婡擻惂屼傪峴偭偰偄傞偲峫偊傜傟傞1丅
KIR偼2偮傑偨偼3偮偺柶塽僌儘僽儕儞條僪儊僀儞傪嵶朎奜僪儊僀儞偲偟偰傕偮type I membrane receptor(偦傟偧傟KIR2D偍傛傃KIR3D偲屇偽傟偰偄傞)偱偁傞丅KIR2D偼僸僩MHC僋儔僗I偺HLA-C傪擣幆偟丄XIR3D偼HLA-A傑偨偼-B傪擣幆偡傞(恾1)丅
KIR2D偼丄戝傑偐偵2庬椶偵暘偗傞偙偲偑偱偒丄HLA-Cw2丆-Cw4丆-Cw5傪娷傓Asn77-Lys80傪傕偮HLA-C傪擣幆偡傞KIR2DL1偲丄HLA-Cw1丆-Cw3丆-Cw7傪娷傓Ser77-Asn80傪傕偮HLA-C傪擣幆偡傞KIR2DL2偲3偱偁傞丅崱夞偼HLA-Cw1丆-Cw3丆-Cw7偵摿堎揑側KIR2DL3偵偮偄偰変乆偑峴偭偨寢徎峔憿夝愅偺榖傪拞怱偵KIR偺MHC僋儔僗 I 偵懳偡傞擣幆婡峔偲偦偺峔憿偵偮偄偰愢柧偟偨偄丅
幚尡偲峫嶡
KIR偺戝検敪尰宯偺峔抸
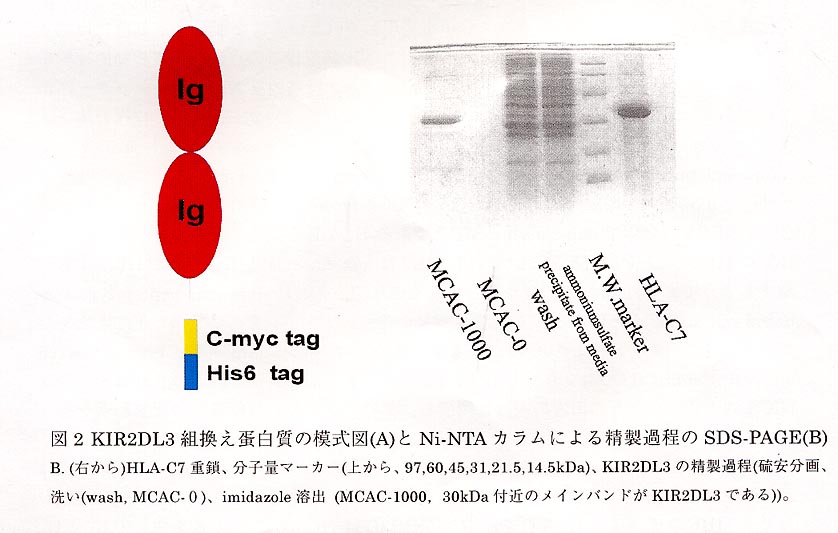
僸僩枛徑寣偺敀寣媴夋暘傪IL2偲PHA偲偲傕偵攟梴偟丄摼傜傟偨嵶朎偐傜mRNA偝傜偵 cDNA傪挷惢偟丄HLA-Cw1丆-Cw3丆-Cw7偵摿堎揑側KIR2DL3偺嵶朎奜僪儊僀儞偺堚揱巕傪揔摉側僾儔僀儅乕傪梡偄偰PCR朄偵傛傝憹暆偟偨丅偙傟傪NcoI媦傃NotI惂尷峺慺僒僀僩傪梡偄偰丄p GEM2儀僋僞乕桼棃偺暘斿敪尰梡儀僋僞乕p KMATH偺T7僾儘儌乕梉乕偺壓棳偵慻傒崬傫偩丅寢壥偲偟偰丄pelB僔僌僫儖傪N枛抂偵帩偪丄C枛抂偵c-myc桼棃偺儕儞僇乕偲His6-tag傪帩偮暘斿敪尰儀僋僞乕傪峔抸偱偒偨(恾2嵍)丅28亷偱攟梴偟丄OD600=0.6-0.8偱0.1mM IPTG偵傛傝敪尰桿摫偟丄偝傜偵堦斢攟梴傪懕偗偨丅慻姺偊抈敀幙偼攟梴忋惔偲儁儕僾儔僘儉夋暘偵暘斿偝傟丄偙傟傪棸埨捑揳偵傛傝夞廂偟偨丅偝傜偵Ni-NTA僇儔儉偲MonoQ(Pharmacia)僇儔儉偵傛傝惛惢偟偨(恾2塃)丅嵟廔廂検偼栺0.5mg/l偱偁偭偨丅
KIR2DL3偺寢徎壔偲X慄僨乕僞應掕
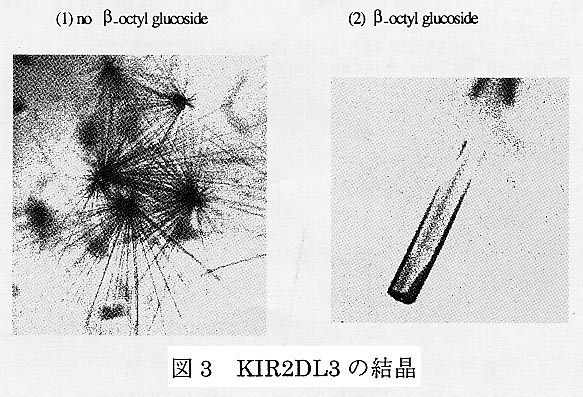
惛惢偟偨抈敀幙傪5-10mg/ml擹搙偵擹弅偟丄寢徎壔忦審傪Crystal Screen(Hampton Research)偱専摙偟偨丅偦偺寢壥丄Crystal Screen II-37(10%PEG8000丆0.1M HEPES pH7.5丆8% Ethylene Glycol)偱恾3嵍偺傛偆側恓忬寢徎偑摼傜傟偨丅偦偙偱丄pH丄墫丄桳婡梟攠側偳傪壛偊偰忦審偺夵椙傪帋傒偨偑丄偆傑偔峴偐側偐偭偨丅偟偐偟丄奅柺妶惈嵻偱偁傞兝-octylglucoside傪0.5%壛偊偨帪偵朹忬寢徎偑摼傜傟偨2(恾3塃)丅X慄僨乕僞應掕傪Photon Factory偺BL6A偱0.05x0.05x0.25mm偺寢徎傪梡偄偰15亷偱峴偭偨丅彫偝偄朹忬寢徎備偊偵懡彮偺堎曽惈偑尒傜傟偨偑丄3侌傑偱偺僨乕僞偑摼傜傟偨丅巜悢晅偗偲僗働乕儕儞僌傪Denzo偲Scalepack偱峴偭偨丅
暘巕抲姺朄偵傛傞夝愅
偡偱偵Wiley傜偺僌儖乕僾偑夞偠僼傽儈儕乕偺KIR2DL1偺寢徎峔憿夝愅3傪敪昞偟偰偄偨偺偱丄偙偺儌僨儖傪梡偄偰丄暘巕抲姺朄偵傛傞夝愅傪帋傒偨丅屻掱傢偐傞傛偆偵KIR偼2偮偺Ig僪儊僀儞偐傜側傝丄偙傟傜偺攝抲偑KIR2DL1偲KIR2DL3偱偐側傝堎側傞偨傔丄暘巕慡懱偱偼暘巕抲姺朄偵傛傞夝愅偑惉岟偟側偐偭偨丅Amore傗X-PLOR傪梡偄偰奺僪儊僀儞偱僒乕僠偟偨偑丄偆傑偔峴偐側偐偭偨丅棟桼偲偟偰峫偊傜傟傞偺偼丄(1)傑偢僨乕僞偺幙偑偁傑傝傛偔側偔丄偐偮暘夝擻偑崅偔側偄偙偲丄偲(2)Ig僪儊僀儞偼lP-sheet偐傜惉傞偨傔丄棤昞偲忋壓偺曽岦偵strand偑怢傃偰偍傝丄偐偮P-sheet偺悈慺寢崌偺棳傟偑偁傞偨傔夝愅傪擄偟偔偟偰偄傞偲峫偊傜傟偨丅偦偙偱Tong偺Combined MolecularReplacement偲偄偆僾儘僌儔儉傪巊偭偨4丅暘巕抲姺朄偱偼捠忢丄寁嶼僗僺乕僪偺愡栺偺偨傔丄傑偢僷僞乕僜儞恾傪傕偲偵暘巕偺夞揮曽埵傪寛傔丄偦偺屻丄暲恑検傪寛傔偰偄偔偲偄偆曽朄傪峴偆丅偙偺僾儘僌儔儉偼偦偺夞揮娭悢偺夝偺寢壥偑捠忢偦傟傎偳傛偔側偄偙偲偑懡偄偙偲傪摜傑偊偰丄偳偺斖埻偺夞揮娭悢偺夝偱傕暲恑偺夝愅偵摉偰偼傔傞偙偲偑偱偒傞傛偆偵嶌傜傟偰偄傞丅傑偨丄偙偺僾儘僌儔儉偼暘巕偺廳側傝傪僠僃僢僋偡傞偙偲偑偱偒傞偺偱丄懡偔偺夝傪僌儔僼傿僢僋僗忋偱妋擣偡傞嶌嬈傪寉尭偡傞偙偲偑偱偒傞丅Tong偺榑暥拞偱傕巜揈偝傟偰偄傞傛偆偵僷僢僉儞僌偺僠僃僢僋偑偐側傝桳岠偱偁傞傛偆偩丅変乆偺KIR偺寢壥傪恾4偵帵偡丅

偙偺傛偆偵奺僪儊僀儞偱夞揮娭悢偺忋埵40%偺斖埻偺夝傪挷傋傞偙偲偵傛傝丄僺乕僋偑偼偭偒傝偲摼傜傟偨丅傑偨丄偙偺応崌F傪梡偄偢偵婯奿壔偝傟偨F傪梡偄傞曽偑傛傝傛偄寢壥傪摫偄偨丅奺僪儊僀儞偺摼傜傟偨夝偺埵抲娭學偑梕堈偵僪儊僀儞栤傪偮側偖偙偲偺偱偒傞埵抲偵偁傝丄惓偟偄夝偑摼傜傟偨偲峫偊傜傟偨丅Rigid-body Refinement傪峴偄丄偦偺屻丄X-PLOR丄Refhlac丄CNS傪梡偄偰惛枾壔傪峴偄丄Rcryst=24.8%(Rfree=32.0%)偺儌僨儖傪峔抸偱偒偨5丅
KIR偺慡懱峔憿偲僪儊僀儞峔憿
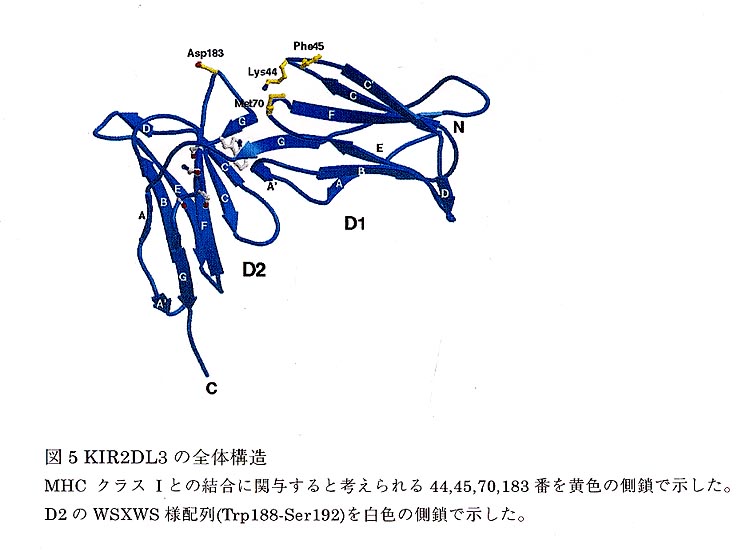
KIR2DL3偺慡懱峔憿偼KIR2DL1偲帡偰丄2偮偺Ig-hke僪儊僀儞(埲壓偱偼N枛懁偺僪儊僀儞傪D1丄C枛懁偺僪儊僀儞傪D2偲屇傇偙偲偵偡傞)偑僞儞僨儉偵暲傫偩峔憿傪偟偰偍傝丄懠偺嵶朎愙拝暘巕傗儗僙僾僞乕偵斾傋偰丄堎忢偵媫側elbow妏傪庢偭偰偄傞(恾5)丅
傑偨丄Sun傜偺僌儖乕僾偑夝愅偟偨丄摨偠偔HLA-Cw1丆-Cw3丆-Cw7偵摿堎揑側傕偆堦偮偺儊儞僶乕KIR2DL26偺峔憿傕帡偰偍傝丄偙傟偼KIR僼傽儈儕乕偺峔憿偺堦斒揑摿挜偲峫偊傜傟偨丅奺僪儊僀儞偼媽棃偺暘椶暘偗偱偼C2set偵帡偰偄傞偺偩偑丄A'strand傪帩偪丄傑偨丄Dstrand偺傛偆側峔憿傕尒傜傟傞偨傔丄尩枾偵偼怴偟偄僞僀僾偺Ig-hke僪儊僀儞偩偲峫偊傜傟偨丅KIR僪儊僀儞傪structural similarity search program Dali (version 2.1)偱夝愅偟偨偲偙傠丄Hematopoietic Receptor(僸僩惉挿儂儖儌儞儗僙僾僞乕傗僒僀僩僇僀儞儗僙僾僞乕偑懏偟偰偄傞)偵娷傑傟傞Fibronectin type III(FN III)僪儊僀儞偵傕帡偰偄傞偑丄傛傝揟宆揑側Ig僪儊僀儞偵帡偰偄偨丅傑偨Hematopoietic Receptor偵摿挜揑偵尒傜傟傞Try-Ser-X-Ser偲偄偆攝楍偺WSXWS儃僢僋僗偵椶帡偟偨Trp-Ser-Asn-Ser-Ser(188-192斣)偲偄偆攝楍偑KIR僪儊僀儞偵傕尒傜傟偨丅偟偐偟丄Hematopoietic Receptor偱偼奺Trp偺朏崄娐偺懁嵔偑摨偠僪儊僀儞撪偺懠偺僗僩儔儞僪偺Arg傗Lys偺懁嵔偲拋彉偩偭偨僷僢僉儞僌傪偟偰偄傞偺偵懳偟偰丄KIR偱偼Trp188偑僪儊僀儞娫偺憡屳嶌梡偵娭梌偟偰偍傝丄慡偔堎側傞峔憿傪帩偭偰偄偨丅
KIR僪儊僀儞偺攝岦偺曄壔
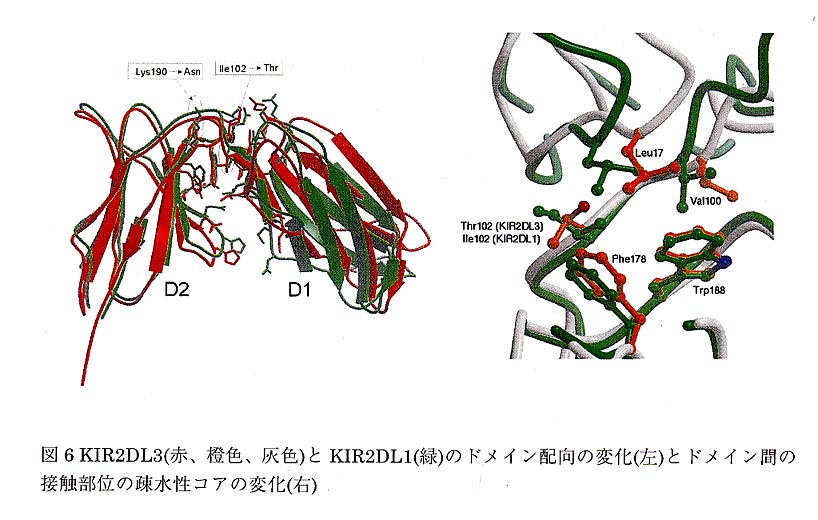
摉弶丄KIR2DL1偺峔憿夝愅偺寢壥偐傜孅嬋偟偨偙偺峔憿偼僪儊僀儞娫偺愙怗晹埵偺峀斖側憡屳嶌梡偐傜棃偰偍傝丄偙偺峔憿偺Flexibility偼掅偄偲峫偊傜傟偨丅偟偐偟丄堄奜偵傕KIR2DL3偺峔憿偱偼KIR2DLl偲斾傋偰丄23搙傕僪儊僀儞偑偢傟偰偄傞偙偲偑傢偐偭偨(恾6)丅偙偺堊丄KIR2DLl偱尒傜傟偨僪儊僀儞栤偺偮側偓栚偐傜墦偄偲偙傠偺憡屳嶌梡偑尒傜傟偢丄偙偺峔憿偼偮側偓栚偵嬤偄晹暘偺憡屳嶌梡偵桼棃偟偰偄傞偲峫偊傜傟偨丅懠偺KIR2DL2偺擇偮偺堎側傞寢徎宯偺峔憿偱5搙傎偳偺僪儊僀儞娫偺偢傟偑尒傜傟偨偑丄媫側elbow妏傪帩偮峔憿偼堐帩偝傟偰偄偨丅傑偨丄偮側偓栚晹暘偺僪儊僀儞栤偺憡屳嶌梡偵娭傢傞傾儈僲巁傪KIR2DL1丆2丆3偱斾妑偟偨偲偙傠丄傎偲傫偳偺傾儈僲巁偑堐帩偝傟偰偄偨偑丄102斣栚偺KIR2DL1偱偼ne102偑丄KIR2DL2丆3偱偼Thr偵曄壔偟偰偄偨丅Thr102偺懁嵔偺悈巁婎偑懚嵼偟偰偄傞偨傔丄廃曈晹偺僪儊僀儞娫偺偮側偓栚晹暘偺慳悈惈僐傾偺攝抲偑曄壔偟偰偄偨(恾6)丅偙偺偙偲偵傛傝KIR2DL2丆3偺峔憿偱偼KIR2DL1傛傝傕彮偟僪儊僀儞栤偺elbow妏偑娚傗偐側偭偰偄傞偺偐傕偟傟側偄丅偙偺傛偆側僪儊僀儞娫偺攝岦偺曄壔偑傑偨KIR2DL1偲KIR2DL2丆3偺儕僈儞僪MHC僋儔僗I偺摿堎惈偵塭嬁偟偰偄傞偙偲傕峫偊傜傟傞丅
MHC僋儔僗I寢崌晹埵偺峔憿
Winter偲Long7偼site-directed mutagenesis偵傛傝KIR2D忋偺MHC僋儔僗I偺婎幙摿堎惈傪寛掕偟偰偄傞晹埵偑丄D1偺CC'儖乕僾偺44斣栚偺巆婎偱偁傞帠傪摨掕偟偨(恾5)丅44斣栚偑Lys偱偁傞KIR2DL2傗3偼HLA-Cw1丆-Cw3, -Cw7傪娷傓僌儖乕僾傪擣幆偟丄懠曽Met偱偁傞KIR2DL1偺応崌偼HLA-Cw2丆-Cw4丆-Cw5傪娷傓僌儖乕僾偵寢崌偡 傞丅偙偺44斣栚巆婎傪岎姺偡傞偩偗偱婎幙偺摿堎惈傪曄姺偡傞偙偲偑偱偒傞丅傑偨丄懠偺僌儖乕僾偺曬崘偐傜丄Dl偺CC乫儖乕僾偺45斣丄D1偺EF儖乕僾偺70-72斣傗D2偺FG儖乕僾偺183斣傕寢崌偵廳梫側巆婎偱偁傞偙偲偑傢偐偭偨丅懠曽丄儕僈儞僪偱偁傞MHC僋儔僗I偺KIR2D寢崌僒僀僩傕摨掕偝傟偰偍傝丄儁僾僠僪寢崌晹埵偺C枛抂懁晹暘(73丆76丆80丆90斣)傪擣幆偡傞8丅摿偵80斣栚偺巆婎偑Lys偱偁傞応崌(HI-Cw4)丄KIR2DL1傪擣幆偟丄Asn偺応崌(HLA-Cw7)丄KIR2DL2傗3傪擣幆偡傞丅偙偺80斣栚巆婎傪岎姺偡傞偩偗偱KIR2D偺摿堎惈傪曄姺偡傞偙偲偑偱偒傞丅
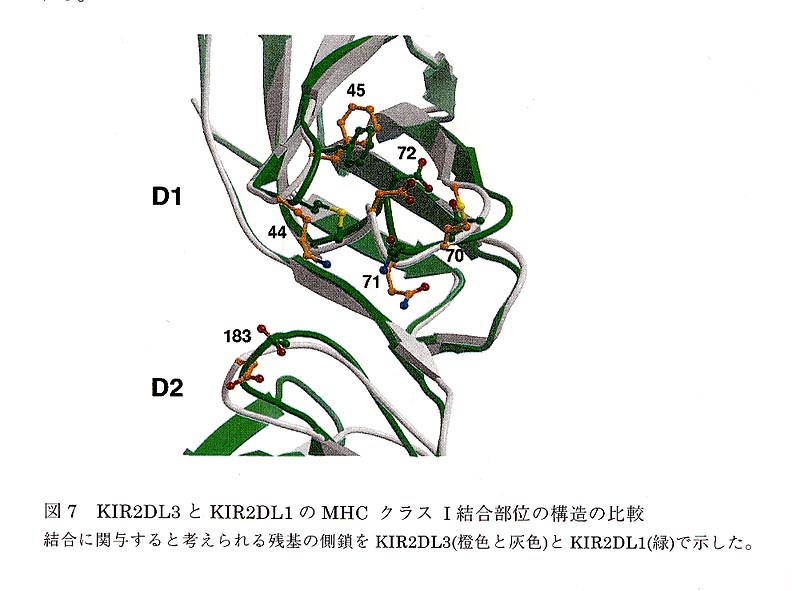
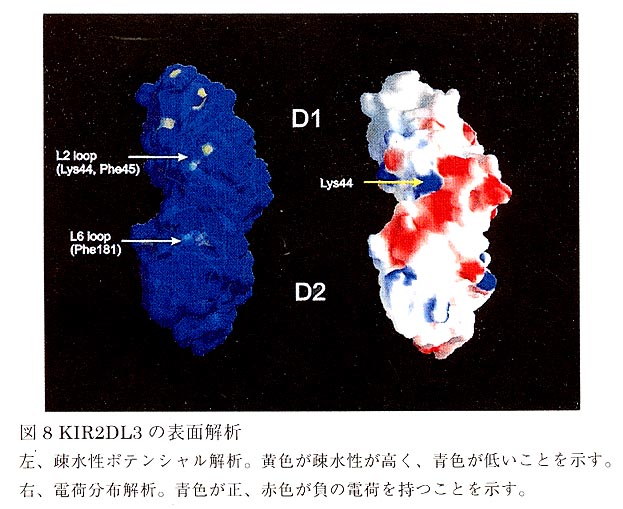
MHC僋儔僗I偺寢崌偡傞偲峫偊傜傟傞晹埵傪KIR2DL1偲KIR2DL3偱斾妑偟偨偲偙傠丄忋偵忋偘偨3偮偺儖乕僾偵偼僪儊僀儞偺摦偒偺塭嬁偑庒姳尒傜傟偨(恾7)丅KIR2DL3偺Lys44(KIR2DLl偱偼Met)偵側偭偨偙偲偵傛傞庡嵔偺戝偒側曄壔偼傎偲傫偳尒傜傟側偐偭偨偑Js44偍傛傃Phe45偺懁嵔偺攝岦偑EF儖乕僾偺曄壔偺塭嬁傪庴偗偰丄KIR2DL1偺偦傟傜偲偼堎側偭偰偄偨丅KIR2DL3偺Pro68(KIR2DL1偱偼Arg)偺偨傔丄D1偺慳悈惈僐傾偺峔惉偑KIR2DL1偲偼堎側傝丄EF儖乕僾偺峔憿曄壔偲摨挷偟偰偄傞傛偆偱偁傞丅傑偨丄Lys44偺懁嵔N兠偑Gln71偲悈慺寢崌偟偰偄傞偑丄抈敀幙偺昞柺揹壸偺夝愅(恾8塃)偐傜丄Lys44偺惓揹壸偑昞柺偵尒傜傟丄寢崌偵塭嬁偡傞偙偲偑峫偊傜傟偨丅傑偨丄hydrophobic potential analysis偺夝愅乹恾8嵍)偐傜丄Lys44偺慳悈惈晹暘偑寢崌晹埵偺拞偱嫮偄慳悈惈傪帵偟偨丅KIR2DL1偲2傪梡偄偨曄堎懱偺夝愅偺寢壥偼Lys44偺惓揹壸偑Lys80傪帩偮HLML-Cw4偺寢崌傪朩偘偰偄傞傛偆偩偑丄扨弮偵揹壸偩偗偱慡偰傪愢柧偱偒側偄偺偱丄慳悈惈晹暘偲偦偺棫懱峔憿傕摿堎惈偍傛傃恊榓惈傪寛掕偟偰偄傞傕偺偲巚傢傟傞丅
KIR2D偲MHC僋儔僗 I 偺憡屳嶌梡偺婡擻夝愅
変乆偼傑偨KIR2D偲悢庬偺MHC僋儔僗I傪梡偄偰丄昞柺僾儔僘儅嫟柭傪梡偄偨寢崌幚尡(BIAcore)傪峴偭偨(慜拠傜丄枹敪昞)丅偦偺寢壥丄KIR2D偲MHC僋儔僗I偺寢崌偺夝棧掕悢偼悢兪M偱偁傝丄偙偺寢崌偼HLA乚C偵傾儘摿堎揑偱偁傝丄偐偮寢崌偟偰偄傞傌僾僠僪偵埶懚偟偰偄偨丅偙偺傛偆側擣幆偼T嵶朎儗僙僾僞乕偺MHC偵懳偡傞擣幆偲帡偰偄傞偑丄kinetics偍傛傃擬椡妛揑僷儔儊乕僞乕偑戝曄堎側偭偰偄偨丅KIR偺MHC偵懳偡傞寢崌偼kinetics偑偲偰傕懍偔丄T嵶朎儗僙僾僞乕偺悢廫攞偱偁傝丄懠偺嵶朎愙拝暘巕偺憡屳嶌梡偲摨偠儗儀儖偱偁偭偨丅傑偨丄擬椡妛揑偵尒傞偲丄KIR偺擣幆偼丄T嵶朎儗僙僾僞乕偺傛偆側戝偒側僄儞僩儘僺乕偺懝幐傪戝偒側僄儞僞儖僺乕偱憡曗偡傞傛偆側宍偱偼側偔丄堦斒揑側抈敀幙亅抈敀幙憡屳嶌梡偵傒傜傟傞傕偺偱偁偭偨丅
崱屻偺揥朷
KIR/ILT儗僙僾僞乕孮偵懏偡傞儊儞僶乕偺拞偵偼NK嵶朎偩偗偱側偔丄寣媴偵暆峀偄敪尰暘晍傪帵偡傕偺偑偁傝丄偙傟傜偺婡峔偑NK嵶朎偩偗偺摿庩側椺偱偼側偔丄峀偔堦斒揑偵柶塽娭楢嵶朎偵傕媦傇傛偆偵側偭偰偒偨丅偙傟傜偺儗僙僾僞乕孮偺夝愅偑恑傓偙偲偵傛傝丄寣媴慡斒偺棟夝偵偮側偑偭偰偄偔偲巚傢傟傞丅傑偨丄KIR/ILT僼傽儈儕乕偺堚揱巕孮偼僸僩愼怓懱19q13.4偵懚嵼偟偰偄傞偑丄Fc兛儗僙僾僞乕(CD89)偺堚揱巕傕摨偠埵抲偵懚嵼偟偰偍傝丄堦師攝楍忋偺憡摨惈傕尒傜傟偨丅嵟嬤夝愅偝傟偨懠偺Fc儗僙僾僞乕(Fc兞儗僙僾僞乕IIb偲Fc8儗僙僾僞乕)偺寢徎峔憿9,10偐傜丄偦傟傜偺2偮偺Ig僪儊僀儞偺攝岦側偳偼KIR偲堎側傞偑丄elbow妏偑媫偱偁傞揰傗儕僈儞僪寢崌晹埵偺埵抲偑KIR偲椶帡偟偰偄傞偙偲偑傢偐偭偨丅偙傟傜偺KIR/ILT僼傽儈儕乕偲Fc儗僙僾僞乕僼傽儈儕乕偲偺娭楢傕嫽枴怺偄丅偄偢傟偺僼傽儈儕乕偲傕儗僙僾僞乕偲儕僈儞僪偺暋崌懱偺夝愅偼側偝傟偰偍傜偢丄崱屻偺壽戣偲側偭偰偄傞丅傑偨丄嵶朎儗儀儖偱偺婡擻偵斀塮偝傟偰偄傞偐傕偟傟側偄丄帺慠柶塽偵娭傢傞KIR偲妉摼柶塽偵娭傢傞T嵶朎儗僙僾僞乕偺MHC僋儔僗I偵懳偡傞擣幆婡峔偺堘偄傕丄偙傟傜偺暋崌懱偺峔憿夝愅偑恑傓偙偲偵傛傝柧傜偐偵側偭偰偔傞偲婜懸偝傟傞丅
偍傢傝偵
KIR敪尰宯偺峔抸偼擔愒拞墰寣塼僙儞僞乕偺廫帤栆晇強挿丄揷強寷帯暃強挿丄壆晹搊巙梇攷巑丄拞嶳婱攷孨丄媏抮埨婓巕偝傫傪偼偠傔偲偡傞懡偔偺僙儞僞乕偺曽乆偲搶嫗戝妛堛妛晹摽塱彑巑嫵庼偺嫤椡傪摼偰妋棫偡傞偙偲偑偱偒偨丅KIR偺寢徎峔憿夝愅偼僆僢僋僗僼僅乕僪戝妛E. Yvonne Jones攷巑偲David I. Stuart嫵庼偲偺嫟摨尋媶偵傛傝峴傢傟丄婡擻夝愅偼僆僢僋僗僼僅乕僪戝妛偺Anton P. van der Merwe攷巑偲偺嫟摨尋媶偱峴傢傟偨丅傑偨丄懡偔偺彆尵傪偄偨偩偒傑偟偨搶杒戝妛岺妛晹孎扟愹嫵庼丄捗杮峗暯攷巑丄憂栻媄弍尋媶強偺徏搰惓柧攷巑丄偍傛傃僆僢僋僗僼僅乕僪戝妛抮悈怣擇攷巑傪偼偠傔偲偡傞Laboratoryof Molecular Biophysics偺曽乆偵姶幱偄偨偟傑偡丅嵟屻偵側傝傑偟偨偑丄Photon Factory(PF)偱偺僨乕僞應掕偵偍偒傑偟偰戝曄偍悽榖偵側傝傑偟偨嶁晹抦暯嫵庼丄搉曈怣媣攷巑傪偼偠傔丄PF偺奆條偵姶幱偄偨偟傑偡丅
恾3偼暥專2偐傜IUCR偺嫋壜傪摼偰揮嵹偟傑偟偨丅
恾5丆6塃丄7偼暥專5偐傜Elsevier Science偺嫋壜傪摼偰揮嵹偟傑偟偨丅
暥專 ( 夝愅偺庤朄丄憰抲丄僾儘僌儔儉偵娭偡傞暥專偼妱垽偝偣偰偄偨偩偒傑偟偨丅)