偼偠傔偵 峔憿惗暔妛揑夝愅朄(X慄寢徎夝愅丆拞惈巕慄寢徎夝愅丆NMR丆揹巕尠旝嬀丆奺庬僾儘乕 僽尠旝嬀側偳)偑栚妎傑偟偄敪揥傪悑偘丆抈敀幙傪巒傔偲偡傞惗懱暘巕偺棫懱峔憿夝愅偑惙傫偵側傝偮偮偁傞丅傕偆堦曽偱丆僸僩傪娷傔偨懡偔偺惗暔偺僎僲儉夝愅偑恑峴偟偮偮偁傞丅偙傟傜偺夝愅朄偲丆挿擭偺楌巎傪傕偮丆抈敀幙岺妛傪娷傔偨抈敀徿婡擻夝愅朄傪婎偵偟偰丆乽偱偒傞偙偲側傜丆栺10枩屄偺堚揱巕偐傜側傞僸僩屄懱慡懱偵偮偄偰丆抈敀幙傪娷傔偨慡惗懱暘巕偺棫懱峔憿傪夝柧偟丆偦傟傜偺棫懱峔憿偵婎偯偄偰暘巕婡擻傪夝愅偟偰丆嵟廔揑偵偼僸僩偺慡惗柦尰徾傪丆暘巕儗儀儖偩偗偱偼側偔丆尨巕儗儀儖偺暘夝擻偱棟夝偟偨偄丅傕偟偦傟偑壜擻偵側傟偽丆堛椕側偳偵傕戝偄偵栶棫偮偺偩偑丆.....乿偲偄偆柌偼丆尰帪揰偺変乆偺妛栤儗儀儖偐傜峫偊傞偲丆摉暘偺娫丆偐側偊傜傟傟偦偆偵側偄丅
側傜偽丆乽僸僩偺堚揱巕栺10枩屄偺拞偱丆偁傜備傞惗暔偵懚嵼偟丆婎杮揑惗柦尰徾偵娭梌偡傞廳梫側堚揱巕栺1000屄傪帩偮1偮偺嵶朎偵偮偄偰偩偗偱傕丆嵶朎慡懱偺惗柦尰徾傪丆傑偢尋媶偟偰偍偒偨偄丅乿偦傟偩偗廳梫側婎杮揑堚揱巕側傜傕偆尋媶偼廔傢偭偰偄偰傕傛偝偦偆偩偑丆幚偼栺113(栺300屄)偺堚揱巕偼丆慡偔婡擻枹抦偱偁傞丅傕偟偦偺300屄偺堚揱巕偺偆偪丆1偮偱傕婡擻偑傢偐傟偽丆戝敪尒偱偁傞丅
偦傟偱偼丆
(1)偦偺傛偆偵廳梫側婎杮揑堚揱巕栺1000屄傪帩偪丆
(2)惗柦尰徾傪棟夝偡傞偨傔偵晄壜寚側"堚揱巕憖嶌宯"偲丆
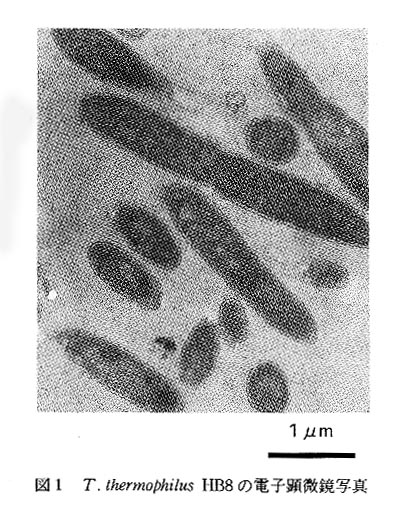
(3)棫懱峔憿夝愅傗暘巕婡擻夝愅偵昁梫側"懴擬惈"偲傪丆寭偹旛偊偨惗暔偼偄傞偺偩傠偆偐?偦偺傛偆側帇揰偱丆偙偺抧媴忋偵1壄庬椶嬤偔懚嵼偡傞偲尵傢傟傞惗暔傪尒夢偟偰傒傞偲丆崅搙岲擬嬠 Thermus thermophilus HB81)(恾12))偑嵟揔偱偁傞偙偲偑傢偐傞丅
偙偺崅搙岲擬嬠偼丆崱偐傜栺30擭慜偵丆擔杮偺埳摛敿搰丒曯壏愹偱丆尰搶嫗栻壢戝妛丒戝搰懽榊嫵庼偵傛偭偰敪尒偝傟偨戝偒偝栺5兪m偺恀惓嵶嬠偱丆85亷偺崅壏偱傕惗堢偱偒傞1)丅
乽峔憿惗暔妛揑夝愅傗婡擻夝愅偵揔偟偨杮崅搙岲擬嬠嵶朎傪儌僨儖惗暔偲偟偰慖傃丆偦偙偵懚嵼偡傞掅暘巕丒崅暘巕偡傋偰偺惗懱暘巕偺婡擻傪棫懱峔憿偵婎偯偄偰夝愅偟偰丆亀崅搙岲擬嬠 Thermus thermophilus HB8 娵偛偲堦旵亁傪尨巕儗儀儖偱棟夝偡傞偙偲乿傪嵟廔栚昗偲偡傞偺偑丆乽崅搙岲擬嬠娵偛偲堦旵僾儘僕僃僋僩乿偱偁傞2)丅偙傟偑恑傔偽丆1偮偺嵶朎慡懱偺惗柦尰徾傪僔儈儐儗乕僔儑儞偡傞偲偄偆柌傊傕嬤偯偔丅
亂1.嵶朎儌僨儖偲偟偰崅搙岲擬嬠(T. thermophilus)傪慖傫偩棟桼亃
偐偮偰丆悽奅拞偺暘巕惗暔妛幰偑丆尋媶懳徾傪戝挵嬠K乚12姅丆偦偺僼傽乕僕傪擖傗T4丆側偳偲寛傔偰奆偱廤拞揑偵尋媶偟丆戝惉岟傪廂傔偨丅杮僾儘僕僃僋僩偺惗暔嵽椏偲偟偰崅搙岲擬嬠(T. thermophilus HB81)))(恾1)傪慖傇棟桼偼丆
1)堚揱巕憖嶌宯偑妋棫偟偨惗暔偺拞偱丆傕偭偲傕崅壏偱惐懅偡傞丅堚揱巕憖嶌偵昁梫側栻嵻懴惈堚揱巕(僇僫儅僀僔儞懴惈堚揱巕)偵偆偄偰偼丆DNA僔儍僢僼儕儞僌朄傪棙梡偟偰懴擬壔偟丆85亷偱傕巊梡壜擻側堚揱巕傪嶌惢偟偨3)丅
2)抈敀幙偺埨掕惈偑崅偔丆寢徎壔傕梕堈側偺偱丆X慄寢徎夝愅丒NMR摍偺峔憿惗暔妛揑
夝愅傗婡擻夝愅偵揔偟偰偄傞丅
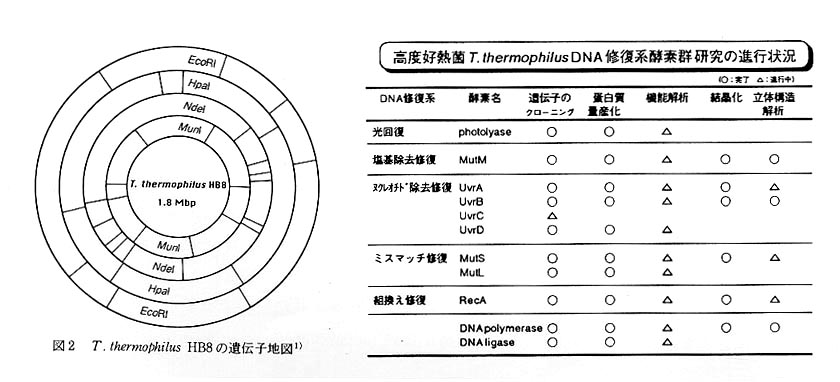
3)僎僲儉僒僀僘偼1.8Mbp(恾22))偲彫偝偔丆嵶朎偺惗柦妶摦偵昁恵側堚揱巕偺傒傪恑壔偺夁掱偱曐帩偟偰偒偨偲峫偊傜傟傞偑丆嵟彮攟抧偱惗堢偡傞偨傔偵昁梫側堚揱巕偼堦懙帩偭偰偄傞丅
4)戝挵嬠偲嫟捠揰偑懡偄偨傔丆戝挵嬠偱妋棫偝傟偨堚揱巕憖嶌宯傕巊梡壜擻偱偁傞丅
5)嵶朎偵偲偭偰婎杮揑偱廳梫側峺慺抈敀幙(椺偊偽丆変乆偺尋媶幒偱夝愅傪恑傔偰偄傞DNA廋暅宯峺慺孮側偳4丆5)(塃壓偺昞))偼丆僸僩側偳偺崅摍摦暔偲傕憡摨惈偑崅偔丆偦偺棫懱峔憿傗婡擻敪尰婡峔偼傎偲傫偳摨偠偱偁傞丅偟偨偑偭偰丆偙偺崅搙岲弉嬠偱夝柧偝傟偨惗柦尰徾偺傎偲傫偳偼丆僸僩傪娷傔偨偁傜備傞惗暔偵嫟捠偱偁傞丅
亂2.杮僾儘僕僃僋僩偺恑峴庤弴亃
1)堚揱巕夝愅(慡僎僲儉偺墫婎攝楍寛掕)
2)奺抈敀徿偺検嶻壔
3)奺抈敀幙偺棫懱峔憿夝愅
4)奺抈敀幙偺婡擻夝愅
5)婡擻枹抦偺抈敀徿偺婡擻夝愅偙傟傜傪丆壜擻側売強偐傜摨帪恑峴偱峴偆丅
1)慡僎僲儉偺墫婎攝楍寛掕
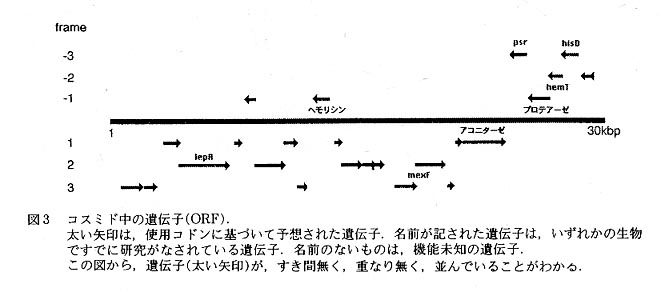
杮崅搙岲擬嬠DNA偼GC娷検偑70%偲崅偄偨傔丆偦偺墫婎攝楍寛掕偼擄偟偄偲峫偊傜傟偨帪戙傕偁偭偨偑丆尰嵼偼壗傜栤戣傪惗偠偰偄側偄丅媡偵丆杮崅搙岲擬嬠偺GC娷検偑崅偄偙偲傪棙梡偡傟偽丆ORF傪梕堈偵悇掕偱偒傞(恾32))丅僎僲儉夝愅偼丆崱擭搙拞偵姰椆偡傞梊掕偱偁傞丅
2)奺抈敀幙偺曮嶻壔
慡抈敀幙丒栺1丆500屄偺偆偪丆偙傟傑偱偵栺300屄偺抈敀幙偺検嶻壔傪庤偑偗偰偄傞丅崅搙岲擬嬠抈敀幙偺検嶻壔偵偼戝挵嬠傪棙梡偡傞丅検嶻壔偺栚埨偼丆1儕僢僩儖偺攟梴塼偐傜丆栺100mg(彮側偔偲傕10mg掱搙)偺惛惢抈敀幙傪摼傞偙偲偱偁傞丅偦偺偨傔偵丆T7僼傽乕僕偺僾儘儌乕僞乕丆傑偨偼丆屚憪嬠偺僞儞僨儉僾儘儌乕僞乕傪帩偮儀僋僞乕傪梡偄偰偄傞丅昁梫偵墳偠偰丆6亊His傑偨偼10亊His偺His乚tag傪枛抂偵晅壛偝偣偨抈敀幙傕検嶻壔偟偰偄傞丅傑偨丆埲慜偼擄偟偄偲峫偊傜傟偰偄偨枌抈敀幙傕丆in vitro 抈敀幙崌惉宯偦偺懠偺曽朄偱検嶻壔偑壜擻側応崌偑偁傞偺偱丆忋婰300屄偺検嶻壔抈敀幙偺拞偵l妱掱搙傪娷傔偰丆検嶻壔傪帋傒偰偄傞丅
偙傟傜検嶻壔僾儔僗儈僪傪丆嬤偄彨棃丆庬乆偺僨乕僞偲嫟偵岞奐偡傞梊掕偱偁傞(偦傟 傑偱偵擖庤傪婓朷偝傟傞応崌偵偼丆屼楢棈壓偝偄(kuramitu@bio.sci.osaka-u.ac.jp))丅
3)奺抈敀幙偺棫懱峔憿夝愅
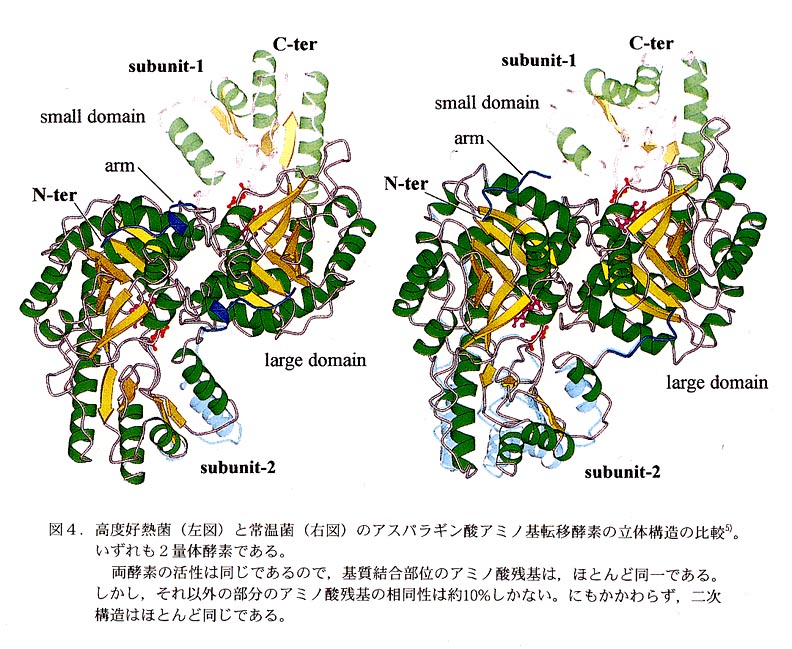
DNA偐傜梊憐偝傟傞抈敀幙偺傾儈僲巁攝楍偺斾妑偩偗偱偼側偔丆棫懱峔憿偺斾妑偑偄偐偵廳梫偐傪幚姶偟偨偺偑丆変乆偺僌儖乕僾偱棫懱峔憿夝愅偟偨傾儈僲婎揮堏峺慺偺椺偱偁傞(恾4)5)丅妶惈晹埵埲奜偺傾儈僲巁攝楍偑傎偲傫偳堎側偭偰偄偰傕丆傎傏摨偠棫懱峔憿傪偲偭偰偄偨丅
偙偺傛偆側棫懱峔憿偺婎杮僷僞乕儞(僼僆乕儖僪)偼丆栺1000庬椶掱搙偟偐側偄偲悇掕偝傟偰偄傞6)(恾5偼偦偺椺)丅偙傟傑偱偵丆栺6妱偑傢偐偭偰偄傞丅奜崙偱偼丆巆傝4妱偺怴婯僼僅乕儖僪傪儕僗僩傾僢僾偡傞偨傔偵丆栺10,000屄偺抈敀幙偺棫懱峔憿傪夝愅偡傞僷僀儘僢僩僾儘僕僃僋僩偑巒傑偭偰偄傞(http://www.nih.gov/nigms/news/meetings/)丅
擔杮偱偼丆桞堦丆乽崅搙岲擬嬠娵偛偲堦旵僽儘僕僃僋僩乿偺抈敀幙偺棫懱峔憿夝愅偺晹暘偩偗偑丆SPring-8偱乽僗僩儔僋僠儐儘乕儉乿尋媶偲偟偰巒傑傞丅
崅搙岲擬嬠T. thermophilus HB8 偺抈敀幙偺棫懱峔憿夝愅偐傜傕丆憡摉悢偺怴婯僼僅乕儖僪偑寛掕偱偒傞偱偁傠偆丅偦偆側傟偽丆抈敀幙偺棫懱峔憿梊應偺惛搙傕忋偑傞偲婜懸偝傟傞丅
4)奺抈敀幙偺暘巕婡擻夝愅
擔杮偺乽崅搙岲擬嬠娵偛偲堦旵僾儘僕僃僋僩乿偺栚昗偼丆傑偢丆崅搙岲擬嬠偺抈敀幙傪 廤拞揑偵棫懱峔憿夝愅偡傞偑丆偦傟偩偗偵偲偳傑傜偢丆寛掕偝傟偨棫懱峔憿偵婎偯偄偰暘巕婡擻夝愅傪峴偄丆嵟廔揑偵偼堦偮偺嵶朎慡懱偺惗柦尰徾傪尨巕儗儀儖偱憤崌揑偵棟夝偟傛偆偲偄偆(Structural and Functional Genomics 偺)憇戝側寁夋偱偁傞丅
偟偐偟丆偙偺暘巕婡擻夝愅偵偮偄偰丆暘巕惗暔妛幰丒峔憿惗暔妛幰偺偄偢傟偐傜傕丆棟夝傪摼傞偙偲偼擄偟偄偺偑尰忬偱偁傞丅
暘巕婡擻夝愅偺岠棪偼丆棫懱峔憿夝愅偑姰椆偡傟偽旘桇揑偵忋徃偡傞丅偦偺暘巕婡擻夝愅偺夁掱偺椺傪恾6偵帵偡丅
峺慺偺暘巕婡擻傪夝愅偡傞嵺偵偼丆傑偢X慄寢徎夝愅傗NMR偺傛偆側峔憿惗暔妛揑夝愅 曽朄偱峺慺暘巕偺棫懱峔憿傪寛掕偟偰偍偔.堦斒偵峺慺斀墳夁掱偼10屄埲忋偺斀墳慺夁掱偐傜側傞偺偱丆棫懱峔憿偐傜摼傜傟傞忣曬偼丆峺慺斀墳慡懱傪棟夝偡傞偨傔偵昁梫側忣曬偺偛偔堦晹偱偁傞丅偦偙偱師偵丆幚嵺偺峺慺斀墳夁掱傪暘岝妛揑曽朄傪棙梡偟偰懍搙榑揑偵夝愅偟丆慗堏忬懺傪娷傔偨斀墳夁掱偺夝愅傪峴偆丅
偝傜偵徻嵶偵丆抈敀幙拞偺奺傾儈僲巁巆婎偺栶妱傪挷傋傞偨傔偵丆堚揱巕憖嶌偱傾儈僲巁巆婎傪抲姺偟偰曄堎宆峺慺傪嶌惢偟丆偦偺棫懱峔憿夝愅偲暘巕婡擻夝愅偲傪峴偆丅偦傟偵傛偭偰丆栰惗宆峺慺拞偵偍偗傞偦偺傾儈僲巁巆婎偺栶妱傪悇掕偡傞(恾6)丅
惗暔壢妛偵偍偄偰偼丆怴婏側尰徾偼娤嶡偝傟偰偄偰傕應掕曽朄傗夝愅曽朄偑妋棫偟偰偄側偄偨傔偵摑堦揑棟夝偑偱偒偰偄側偄彅尰徾偑悢懡偔巆偝傟偰偄傞丅偦傟傜傪棟夝偡傞偨傔偵傕丆怴婯側暘巕婡擻夝愅曽朄偺奐敪傪惛椡揑偵恑傔傞昁梫偑偁傞丅
5)婡擻枹抦偺抈敀幙偺暘巕婡擻夝愅
僠僢僾僥僋僲儘僕乕丆壏搙姶庴惈堚揱巕攋夡姅丆抈敀幙偺擇師尦揹婥塲摦丆棫懱峔憿夝愅丆偦偺懠尰嵼峫偊摼傞偁傜備傞曽朄傪慻傒崌傢偣傞丅
婡擻枹抦偺棫懱峔憿夝愅傪峴偭偰傕丆偙傟傑偱偼嬈愌偲偟偰傎偲傫偳擣傔傜傟側偐偭偨丅偟偐偟丆偨偲偊婡擻枹抦偱傕丆懠偺惗暔偵嫟捠偺儂儌儘僌偑尒偄弌偝傟傞傛偆側応崌偵偼丆偦偺棫懱峔憿夝愅偵傛偭偰丆戝敪尒偺偛偔庤慜傑偱摓払偟偰偄傞壜擻惈偑偁傞丅
岾偄丆偙偺傛偆側婡擻枹抦抈敀幙偺棫懱峔憿夝愅寢壥傪宖嵹偡傞嶨帍偑娫傕側偔敪姧偝傟傞傛偆偱偁傞丅
6)嵶朎慡懱偺惗柦尰徾偺棟夝傪栚巜偟偰
慜弎偺傛偆偵偟偰丆屄乆偺暘巕婡擻傪夝柧偟偨屻丆暋崌宯惗懱暘巕偺夝愅傪峴偄丆傛傝崅師偺惗柦妶摦偺夝愅傪峴偆丅
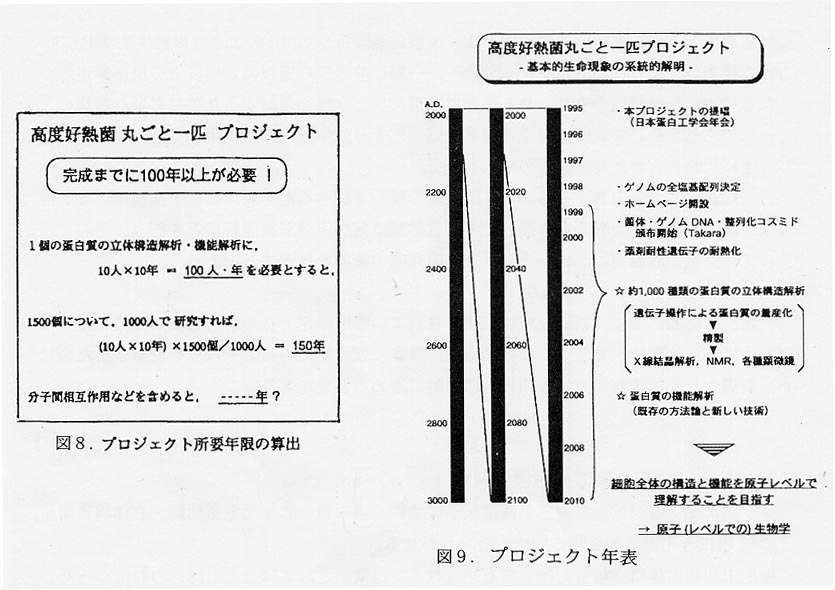
変乆偺宱尡偵傛傟偽丆堦偮偺峺慺抈敀幙偵偮偄偰棫懱峔憿偐傜暘巕婡擻傑偱傪揙掙偟偰夝愅偟傛偆偲偡傟偽丆10恖偺尋媶幰偱彮側偔偲傕10擭栤偼梫偡傞丅杮崅搙岲擬嬠偵偼栺1500庬椶偺抈敀幙偑偁傞偺偱7)(恾7)丆崱屻夝愅曽朄偺旘桇揑敪揥偑柍偗傟偽丆堦捠傝尋媶偡傞偩偗偱傕丆1000恖偺尋媶幰偱100擭埲忋傪梫偡傞丅偝傜偵丆偦傟傜暋崌宯偺憡屳嶌梡傑偱傕夝愅偟傛偆偲偡傞偲丆婥偑墦偔側傞傛偆側擭寧傪梫偡傞(恾8丆9)丅偟偐偟丆偦偺尋媶夁掱偱摼傜傟傞惉壥偵偼偐傝抦傟側偄傕偺偑偁傞偙偲偼丆梕堈偵憐憸偱偒傞丅
偙偺僾儘僕僃僋僩慡懱偼丆惗柦壢妛偺婎慴尋媶暘栰偵偍偗傞憇戝偱偁傞丅杮婥偱庢傝慻傔偽丆尰嵼偺擼壢妛傗寁嶼婡壢妛埲忋偺僾儘僕僃僋僩偲側傞丅
岾偄丆崅搙岲擬嬠T. thermophilus HB8偺僎僲儉夝愅丒抈敀幙偺検嶻壔丒抈敀幙偺棫懱峔憿夝愅偩偗偵偮偄偰偼丆棟尋偺乽僗僩儔僋僠儐儘乕儉乿楢実尋媶傪妀偵偟偰丆巒傔傞偙偲偑壜擻偵側偭偨丅偦偺棫懱峔憿夝愅寢壥偵婎偯偄偰暘巕婡擻夝愅偑恑傒丆堦偮偺嵶朎慡懱偺惗柦尰徾偺棟夝傊彮偟偱傕嬤偢偔偺偑柌偱偁傞丅
偍傢傝偵
嵟嬤偱偼庬乆偺惗暔偺堚揱巕偑師乆偲寛傔傜傟丆婡擻枹抦堚揱巕偺婡擻夝柧偑帋傒傜傟偰偄傞偑抈敀幙偺埨掕惈偑掅偐偭偨傝丆抈敀幙偺寢徎壔偑擄偟偄偨傔偵丆昁偢偟傕惉岟偟偰偄側偄丅傑偨丆懴擬惗暔偱偁偭偰傕堚揱巕憖嶌宯偑柍偄偙偲偑偁傞丅嵟弶偵傕弎傋偨傛偆偵丆抈敀幙傗DNA暘巕偺棫懱峔憿偵婎偯偄偰嵶朎慡懱傪棟夝偟傛偆偲偡傞側傜偽丆堚揱巕憖嶌宯偑妋棫偟偨惗暔偺拞偱丆傕偭偲傕懴擬惈偑崅偄傕偺(T. thermophilus HB8)傪慖傇偺偑摼嶔偲峫偊傜傟傞丅偝傜偵偙偺崅搙岲擬嬠偺傛偆偵彫偝側僎僲儉偵巆偝傟偨堚揱巕偼丆偁傜備傞惗暔偵昁恵偱嫟捠側堚揱巕傪帩偭偰偄傞妋棪偑崅偄丅偟偨偑偭偰丆峔憿惗暔妛揑夝愅朄偦偺懠偺夝愅朄傪嬱巊偟偰偙偺崅搙岲擬嬠傪宯摑揑偵尋媶偡傞偙偲偼丆婎杮揑惗柦尰徾傪憤崌揑偵棟夝偡傞偨傔偺嬤摴偱偁傞偲峫偊傜傟傞丅
偨偐偑堦旵偺彫偝側崅搙岲擬嬠偩偑丆偦偺僾儘僕僃僋僩偺憇戝偝偵婥晅偄偨帪丆乽偙偺僾儘僕僄僋僩偼丆彮悢僌儖乕僾偺偨傔偺尋媶偱偼側偔丆枹棃偺恖椶偺偨傔偺儃儔儞僥僀傾揑婎慴尋媶偱偁傞丅偙偺楢実尋媶偺惉斲偼丆偄偐偵尋媶幰偑嫤椡偟崌偊傞偐偵偐偐偭偰偄傞丅乿偲巚偆偲摨帪偵丆嶁晹僾儘僕僃僋僩偺專恎揑塣塩曽恓傪庤杮偵偟偨偄偲巚偭偰偄傞丅
亂嶲峫暥專亃